Introduction
The Asian soybean rust, Phakopsora pachyrhizi, has been known to drastically reduce yields in Asia. In areas where the pathogen occurs commonly, yield losses up to 80% have been reported. The pathogen has been limited to the Eastern hemisphere until it was found in Hawaii in 1994 (Fig. 1). Currently, the distribution of P. pachyrhizi includes Africa, Asia, Australia, South America, and Hawaii. The rapid spread of P. pachyrhizi and potential for severe yield losses makes this the most destructive foliar disease of soybean. Soybean rust could have a major impact on both total soybean production and production costs in the U.S.
| |
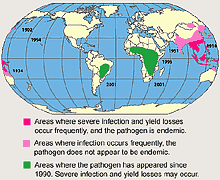
Fig. 1. Distribution of soybean rust caused by Phakopsora pachyrhizi. |
|
In general, conditions that promote good growth and full canopy development of the soybean crop are also the conditions most suitable for development of soybean rust. Temperatures in the range of 15 to 28°C are optimal for infection and spread of the pathogen as long as there is at least 6 to 12 hours of moisture (32). Moisture, as rain or dew, promotes the spread of the pathogen once infection has taken place. The normal temperatures and moistures found in the North Central region of the U.S. are well within the ranges that would result in severe yield losses. In 1984, an economic risk analysis projected that the potential losses in the U.S. would be $7.1 billion per year, once soybean rust was established in the main soybean growing area of the U.S. (24). A conservative prediction indicated yield losses greater than 10% in nearly all the U.S. soybean growing areas with losses up to 50% in the Mississippi delta and southeastern costal states (50).
Geographic distribution
The first report of the disease was from Japan in 1902 (Fig. 1). By 1934 the pathogen had been found in several other Asian countries and as far south as Australia (8). In India, soybean rust was first reported on soybean in 1951 (42). There have been several early reports of soybean rust in equatorial Africa (5,19), but the first confirmed report of P. pachyrhizi on the African continent was in 1996 from Kenya, Rwanda, and Uganda. Since then, the pathogen has spread south with reports from Zambia and Zimbabwe in 1998, Mozambique in 2000 and South Africa in 2001 (25). The westward movement of the pathogen on the African continent was reported from Nigeria in 1999 (3). The disease was found in Hawaii in 1994 on cultivated soybeans on the islands of Oahu, Kakaha, Kauai, and Hilo (20). With this introduction into the U.S., a workshop was organized to discuss issues related to the possible introduction of soybean rust into the continental U.S. (45).
The first detection of P. pachyrhizi in the new world was in Paraguay in February of 2001. The disease was found on soybeans in a limited number of fields in the Parana River basin bordering Brazil. By 2002, soybean rust was widespread throughout Paraguay and in limited areas of Brazil bordering Paraguay, with reports of severe disease in some fields in both countries (33). The pathogen also was found in a limited area in northern Argentina (39). During the 2003 growing season, the pathogen was detected in most of the soybean growing regions of Brazil with a conservative yield loss estimate of 2.2 MMT, or approximately 5% of the annual production. In Paraguay, yield loses from rust were limited due to dry conditions, while in Argentina the disease did not spread to the major production areas. The continental U.S. is the only major soybean producing area in the world where soybean rust has not been found.
Surveillance and Identification
With the confirmed identification of the Asian soybean rust in South America, the U.S. Department of Agriculture’s Animal Plant Health Inspection Service (APHIS), Agricultural Research Service (ARS), Cooperative State Education and Extension Service (CSREES), the United Soybean Board (USB), and other federal and state agencies are concerned about the introduction of soybean rust in the continental U.S. Other plant pathogens have spread from South and Central America into the U.S. with barley stripe rust and sorghum ergot among the latest examples. There are major efforts in place for surveillance, detection, and diagnosis of soybean rust by USDA-APHIS in conjunction with the CSREES Regional Plant Disease Diagnostic Centers. APHIS has developed a National Strategic Action Plan for dealing with soybean rust. If soybean rust is suspected to be present on soybean or other hosts in the continental U.S., samples are to be submitted to the plant disease diagnostic laboratory at state universities for identification.
Two Phakopsora species are known to cause soybean rust (36). The more aggressive species is P. pachyrhizi, known as the Asian soybean rust. Phakopsora meibomiae, the less virulent species, has only been found limited areas in the Western hemisphere, and it is not known to cause severe yield losses in soybean. The focus of the recent concern, most of the literature, and this paper is on P. pachyrhizi. Scientists at the USDA-ARS Foreign Disease-Weed Science Research Unit (FDWSRU) (13) have developed a molecular tool using PCR primers that provides rapid and accurate identification of P. pachyrhizi and P. meibomiae. This assay and the identification by the USDA-APHIS National Mycologist will provide the official confirmation that P. pachyrhizi has entered the continental U.S.
Host Range
Phakopsora pachyrhizi infects over 95 species of plants from more than 42 genera (6), including soybean and related Glycine species (36,40,47). Included in the list are many of the wild and edible legumes. The full host range of both rust species has not been clearly identified and may be complicated by pathotypes and differential reactions within host species (10,11). Included in the list is Kudzu (Pueraria lobata), which is widespread in the U.S. and could serve as an inoculum reservoir or bridge host for P. pachyrhizi in the southern U.S. (Fig. 2). Kudzu, growing along roadsides and in ditch banks in Brazil and Paraguay, was severely infected with rust and showed no apparent loss of plant vigor (M. Miles, pers. obs.). The broad host range of the two species of Phakopsora that infect soybean is unusual among rust pathogens, as most rust species have a narrow host range that is limited to a few plant species. The large number of host species increases the likelihood that this pathogen will survive and over winter in the southern U.S., as well as in Central America.
| |

E
|
|

F
|
|
| |

G |
|
Fig. 2. (A) Phakopsora pachyrhizi lesions on the upper surface of a soybean leaf, (B) lesions on a perennial Glycine species showing a resistance response, (C) lesions on Kudzu growing along side fields in Paraguay, (D) symptoms on infected soybean plants 9 days after inoculation, (E) lesions on a susceptible red kidney bean, (F) heavily infected greenhouse grown soybean plants, and (G) soybean leaf with older lesions; note the darker color of the lesion.
Signs and Symptoms
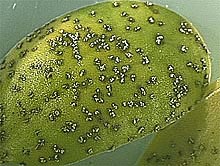
The most common symptoms of soybean rust on cultivated soybean are 2 to 5 mm2 tan to dark brown polygonal lesions. Within each of the lesions are one to many erumpent, globose uredinia (Fig. 3). Urediniospores are released through the circular ostiole. Lesions are found on petioles, pods, and stems but are most abundant on leaves (Fig. 4). As rust severity increases, premature defoliation and early maturation of plants is common.
Fig. 3. (A) Early infection by Phakopsora pachyrhizi on a susceptible soybean, (B) susceptible (TAN) lesions, (C and D) red brown (RB type) lesion with some sporulation, (E) RB lesion with circular ostiole and spores, and (F) RB lesion with no sporulation.

A
|
|
B
|
| |

C
|
|

D
|
|
| |

E |
|
Fig. 4. (A) Soybean rust pustules caused by Phakopsora pachyrhizi on the upper side of a soybean leaf, (B) on a soybean cotyledon, (C) on petioles, (D) stems, and (E) on the under side of a soybean leaf.
Life cycle and infection process
Two spore types are known in P. pachyrhizi. The urediniospore is the common spore type found throughout the season. The abundantly produced urediniospores are readily wind dispersed and multiple spore cycles occur throughout the season. Telia and teliospores have been found on infected plants late in the season in Asia as well as in the greenhouse (6,51). Under laboratory conditions, teliospores have been germinated and basidiospores produced (41). Since no alternate host has been identified, there has been no further characterization of the life cycle.
The infection process starts when urediniospores germinate to produce a single germ tube that grows across the leaf surface, 5 to 400 µm, until an appressorium forms. Appressoria form over anticlinal walls or over the center of epidermal cells, but rarely over stomata (4,22,31). Penetration of epidermal cells is by direct penetration through the cuticle by an appressorial peg. When appressoria form over stomata, the hyphae penetrate one of the guard cells rather than entering the leaf through the stomatal opening. This rust and related species are unique in their ability to directly penetrate the epidermis; most rust pathogens enter the leaf through stomatal openings and penetrate cells once inside the leaf. The direct penetration of the epidermal cells and the non-specific induction of appressoria (21) in the infection process of P. pachyrhizi may aid in understanding the broad host range of the pathogen and may have consequences in the development of resistant cultivars.
Uredinia can develop 5 to 8 days after infection by urediniospores. The first urediniospores can be produced as early as 9 days after infection, and spore production can continue for up to 3 weeks (22,27). Uredinia may develop for up to 4 weeks after a single inoculation, and secondary uredinia will arise on the margins of the initial infections for an additional 8 weeks. Thus, from an initial infection, there could be first generation pustules that maintain sporulation for up to 15 weeks. Even under dry conditions this extended sporulation capacity allows the pathogen to persist and remain a threat. If conditions for re-infection are sporadic throughout the season, significant inoculum potential still remains from the initial infection to reestablish an epidemic. Successful infection is dependant on the availability of moisture on plant surfaces. At least 6 hours of free moisture is needed for infection with maximum infections occurring with 10 to 12 hours of free moisture. Temperatures between 15 and 28°C are ideal for infection (12,16,28,32).
Management Recommendations
Most research on control has focused on the use of fungicides and host plant resistance. Some cultural practices have been recommended that minimize the impact of rust (12,16). The recommendations differed, but were based upon avoiding the conditions that promote disease development or were practices that optimized overall yields. Research on biological control has been limited.
Host plant resistance was first reported in the 1960s from field evaluations in Taiwan. Physiological races of P. pachyrhizi were first described in 1966 when a set of nine single urediniospore isolates were inoculated onto six soybean and five legume accessions (26). The reactions of the nine isolates were similar on all six of the soybean genotypes, but six pathotypes were identified based upon their reactions on the legume accessions. The first example of virulence diversity on soybean cultivars was described in Queensland, Australia (29) where one rust isolate was found to be virulent on the cultivar ‘Wills’ but avirulent on the accession PI 200492, while another isolate was virulent on both soybean genotypes. Several other studies have also shown considerable variation in virulence among isolates from the same field as well as isolates collected from wide geographical areas (1,38,43).
Specific resistance to P. pachyrhizi is known, and four single dominant genes have been identified as Rpp1(30), Rpp2 (8), Rpp3 (7,8,18), and Rpp4 (17). These four genes condition resistance to a limited set of rust isolates (Table 1). Rpp1 was described as having an immune reaction when inoculated with a few isolates, including India 73-1. Inoculation of most rust isolates on plants containing Rpp1 or the other genes, produce a resistant red-brown (RB) lesion with no or sparsely sporulating uredinia. The RB lesion type is considered to be a resistant lesion type when compared to a fully susceptible TAN lesion (Fig. 3). Single gene resistance has not been durable, and the usefulness of the single genes was lost soon after the sources were identified (23). For example, the accession PI230970 was identified as resistant in field evaluations in 1971-1973. In 1976, a few susceptible lesions were observed on plants in the field; by 1978, most of the lesions were of the susceptible TAN type (6). Similarly, Komata was identified in germplasm evaluations done during 1961-1963 (6). By 1966, susceptible lesions were found on plants of Komata in field trails, and by the mid 1970’s the line was not considered to be a useful source of resistance (23). The resistance in Ankur, identified in the early 1970’s (46) was overcome in the early 1980’s (6), and provided another example where a single gene for resistance was not useful. Only Bing Nang, the source of the Rpp4 gene, has not been reported to be defeated in the field, although greenhouse inoculations have identified P. pachyrhizi isolates that are virulent on it (Frederick et al., unpublished).
Table 1. Named single genes, original sources and Phakopsora pachyrhizi isolates used in studies of the inheritance of resistance to soybean rust
Named single
gene |
Accession number
and cultivar name
of original source |
Phakopsora pachyrhizi isolatesa |
| Resistant reaction |
Susceptible reaction |
| Rpp1 |
PI200492
Komata |
IN 73-1bc |
TW 72-1,
TW 80-2
(18,30)d |
| Rpp2 |
PI230970 |
AU 72-1c,
IN 73-1c,
PH 77-1c,
TW 72-1c |
TW 80-2
(7,18,30)d |
| Rpp3 |
PI462312
Ankur |
IN 73-1c |
TW 72-1,
TW 80-2
(18)d |
| Rpp4 |
PI459025
Bing Nang |
IN 73-1c,
TW 72-1c,
TW 80-2c |
(17)d |
a AU = Australia, IN = India, PH = Philippines, TW = Taiwan.
b Immune reaction type.
c Isolates used in original inheritance studies to examine segregation patterns.
d Reference citation.
Partial resistance, or rate reducing resistance, is also known in soybean (49). Lines with partial resistance in field evaluations were rated as moderately resistant, since fewer lesions developed on plants throughout the season. In greenhouse studies, host-pathogen combinations that resulted in RB reaction types tended to have longer latent periods, lower rates of increase in pustule number over time, and smaller lesions compared with susceptible interactions that resulted in a TAN reaction type (9,27). Identification and utilization of partial resistance in breeding programs has been limited. The evaluation methods are time consuming and difficult to incorporate into breeding programs and have been limited to use with advanced generations. These difficulties, at least in part, lead to the development of a strategy to select genotypes with tolerance (2,14,49).
Tolerance is the strategy to select genotypes with high yield potential that have less yield loss from soybean rust. Screening for tolerance to soybean rust was started at the Asian Vegetable Research and Development Center (2,14), where yields from paired plots, with and without the fungicide Dithane M-45 applied every 2 weeks, were compared to determine losses due to rust. High yielding cultivars with less yield loss under severe rust conditions were considered to be tolerant. Rust development rates and estimates of rust severity on foliage were not correlated with yield loss in tolerant materials. Using fungicide protected plots as yield checks, tolerant lines from breeding populations were identified as early as the F5 generation without having to take detailed notes on rust severity (1,14).
Most of the work on chemical control of soybean rust has been done in Asia with more recent information from Africa and South America. The fungicide Dithane M-45 was one of the earliest fungicides reported to control soybean rust (16,44). Since the compound is a protectant, it requires frequent applications (7 to 10 days) and is removed by rain. Several reports on yield protection from Dithane M-45 were inconclusive (34,44). Other compounds have been evaluated and found to be effective in controlling the pathogen (Table 2). The simplest method for determining the need to start fungicide applications to control soybean rust was presented in the Taiwanese grower’s journal ‘Harvest’. The recommendation was to harvest leaves in the lowest part of the canopy weekly, starting three weeks after germination. The leaves were placed in plastic bags and left in a shaded area. Growers were advised to start fungicide applications when these leaves develop rust pustules (16). The most current recommendations for chemical control have come from Zimbabwe and South Africa (C. Levy, pers. comm.). Three fungicide applications were recommended, with the first application 50 days after planting or at early flowering. Second and third applications were 20 days apart, at 70 and 90 days after planting, to provide protection during pod fill. In low incidence areas two applications were recommended to prevent early leaf drop and yield loss. Punch (flusilazole/carbendazim) and Shavit (triadimenol) were found to be the most effective compounds for controlling soybean rust.
Table 2. Partial list of fungicides used to control soybean rust caused by Phakopsora pachyrhizi, with a summary of effectiveness and recommendations.
Active
ingredient |
Compound |
Country
reported |
Summary of
application trials and
recommendations
in the literature |
Referencesb |
| Triadimefona |
Bayleton |
Thailand, India, Taiwan, Japan, Philippines |
Protection was inconsistent when compared to Dithane M45, although it was used as a control in yield loss studies. EBDC fungicides appear to be more effective, but in limited testing increased yield up to 33%. First application at flowering, 10 to 20 day intervals. |
(16,37,48) |
| Thiabendazole |
Benlate
Topsin |
Thailand
Zimbabwe South Africa |
Off registration in U.S., not as effective as Dithane M45, was found to be effective only when used with Plantvax. Phytotoxic as a seed treatment. Topsin M registered for use on soybeans in the U.S. |
(16) (C. Levy, pers. comm.) |
| Chlorothaninol |
Bravo
Equus |
South America
India |
Limited data available. Yield protection similar to or less than Mancozeb. Not as effective as other compounds in some studies. Registered for use on soybeans in the U.S. |
(37,48) |
Ethlenebisdithio-
carbamates
(EBDC)a |
Dithane M45
Mancozeb Manzate D
Zineb Maneb |
Australia, China, India Philippines, Taiwan |
The EBDC products have been effective in controlling soybean rust when applied 7 to 21 days apart, with the first applications as early as three weeks after planting and as late as flowering. Not all studies showed control of yield increases. |
(1,2,16,35,44) |
| Oxycarboxina |
Plantvax |
India
Taiwan Thailand |
Not as effective as Dithane M45 or Manzate D, did not always control rust, yield protection varied by study. Apply when lesions first appear, then at 7-day intervals. |
(4) |
| Hexaconazolea |
Contof |
India |
Effective in reducing disease and protecting yield, 25% yield increase in limited study. |
(37) |
| Propiconazolea |
Tilt |
India
Brazil |
Effective in reducing disease and protecting yield, 33% yield increase in limited studies. Two applications, 15 days apart, starting at flowering. |
(37) |
| Difenconazole |
Score |
Zimbabwe
South Africa India |
Yield protection varied by study, more effective than Mancozeb. Two or three applications needed, starting at flowering. |
(37) (C. Levy, pers. comm.) |
| Tridimefona |
Shavit |
Zimbabwe
South Africa India |
Extremely effective in reducing disease incidence. Highest yielding treatment. Two or three applications needed, starting at flowering. |
(37) |
Flusilazole/
Carbendazima |
Punch Extra |
Zimbabwe
South Africa |
One of most effective fungicides in Africa. Two or three applications needed, starting at flowering. |
(C. Levy, pers. comm.) |
| Tebuconazolea |
Folicure |
Zimbabwe
South Africa |
Not effective in limited testing in Africa. |
(C. Levy, pers. comm.) |
| Azoxystrobin |
Quadris |
Brazil |
Limited data. Good control but single, late application did not control rust or protect yield. Registered for use on soybean in the U.S. |
(A. Talley, pers. comm.) |
a Not registered on soybean in U.S.
b For additional summaries of references see record numbers 182, 185, 285, 320, 407, and 408 in (16).
There are only a few fungicide compounds currently registered for foliar application on soybean in the U.S. These are Bravo and Equus (chlorothaninol), Benlate and Topsin M (thiabendazole) and Quadris (azoxystrobularin). Efficacy data is limited on all three classes of compounds. Chlorothaninol and thiabendazole have been reported to be as effective or less effective than Mancozeb or Dithane M45 (16,37,48). Quadris, an azoxystrobularin, has limited data from efficacy trials in Brazil and Africa but does control soybean rust. Additional fungicides are needed for economic reasons as well as for resistance management strategies.
There are still many questions to be answered regarding the use of chemicals to control soybean rust. Most soybeans are not treated with foliar fungicides, so there is little known about application methods or the effect of fungicides on the crop. Current fungicide application methods may not be sufficient to control the pathogen. Early in the season the pathogen is found on the lower leaves of the plant where lesion numbers increase and inoculum builds up. As the plants start to flower this build up of inoculum increases and the infection moves up the plant as the lower leaves die and drop off. To get maximum yields, the crop needs protection from flowering through pod fill. During this period the plant canopy is very dense, and can be an effective barrier to penetration from fungicides applied over the top of the canopy. Most of the available fungicides are not systemic, or if systemic will not move down in the plant. Application methods need to be developed that will deliver enough fungicide, uniformly, into the lower portion of the soybean canopy after flowering. The economics of fungicide application, timing, number of sprays, and which fungicides to use for the most cost effective control all need to be addressed by additional research.
Current Research
Since the report of soybean rust in Hawaii in 1994, the USDA-ARS has renewed its support for soybean rust research in the U.S. The FDWSRU at Ft. Detrick is the focal point of this research with additional collaborators in the USDA and states universities with additional support from the United Soybean Board. Part of the research focus has been to identify resistant germplasm and evaluate fungicide efficacy. Other research priorities include genome sequencing projects of both soybean rust pathogens, P. pachyrhizi and P. meibomiae, in collaboration with the DOE Joint Genome Institute, developing methods for molecular identification of rust isolates, and monitoring gene expression of both pathogen and host during susceptible and immune reactions. In addition, ARS is funding a project with U.S. and international collaborators to develop remote sensing techniques for monitoring the spread of soybean rust within fields.
There are over 16,000 soybean accessions in the USDA Germplasm Collection located at the University of Illinois. These soybean accessions are being evaluated for resistance to P. pachyrhizi in the USDA-ARS FDWSRU Biosafety Level 3 Containment Greenhouses at Fort Detrick, MD, along with commercial and public cultivars grown in the U.S. (Fig. 5). The germplasm evaluations are done on seedlings using a mixture of isolates from Africa, Asia and South America. None of the U.S. commercial cultivars evaluated were found to be resistant to the mixed inoculum, but some differences were observed in levels of susceptibility. Some cultivars were moderately susceptible when compared to other cultivars that were susceptible and super susceptible. From 3,600 soybean accessions screened to date, less than 100 have been identified as having some level of resistance. These soybean accessions will be further evaluated using individual isolates to detect race specific and/or partial resistance. They also will be planted in field trials in Brazil, Paraguay, China, Thailand, South Africa and Zimbabwe to be evaluated for adult plant resistance (Fig. 6). Additionally, the 1,000 plus G. soja accessions will be screened along with verification of the perennial Glycine spp. previously reported as having resistance (15). As sources of resistance are identified, crosses will be made to incorporate this resistance into adapted backgrounds.
 |
|
 |
Fig. 5. Flats of soybean accessions being evaluated for reaction to Phakopsora pachyrhizi in the USDA-Agricultural Research Service, Foreign Disease-Weed Science Research Unit Biosafety Level-3 Containment Greenhouse at Ft. Detrick, MD (left), and a close up of soybean plants 14 days after inoculation (right).
Fig. 6. Adult plants being screened for Phakopsora pachyrhizi reactions in a screen house at the Centro Regional de Investigación Agrícola, one of the research stations for the Ministry of Agriculture and Livestock in Paraguay (left, courtesy of Ing. Wilfrido Morel Paiva., CRIA), and adult plants being screened for soybean rust reaction in a greenhouse at EMBRAPA soja (The Brazilian Agricultural Research Corporation Soybean Unit) in Brazil (right, courtesy of Dr. José Tadashi Yorinori, EMBRAPA Soja).
Fungicide applications will be the primary tool used to control soybean rust should it arrive in the U.S. in the near future. Our current research is designed to answer some of the questions raised about fungicide application and efficacy on soybean. The first question to be addressed is which fungicides, among those that could be or are already registered for use in the U.S., are effective in controlling soybean rust. To answer this question a multi-location efficacy trial has been established to evaluate 17 fungicides in a replicated trial where two vs. three applications were compared (Fig. 7). The second question to be addressed involves improving the methods of fungicide application. A multi-state project, funded by the Critical Issues Program of USDA-CSREES, to examine aerial and ground application methods is in progress. The aerial application experiment will evaluate canopy penetration using different volumes of water with two fungicides. The ground application experiment will compare canopy penetration and coverage with different application volumes and nozzle types, within and above the canopy. Both ground and aerial applications will start at flowering with additional applications at 20-day intervals. Crucial questions still remain regarding the availability of fungicides and the equipment to apply them should a severe epidemic occur in the U.S. There is a further need to determine the timing, minimum number of applications, and effective rates for each compound, as well as the economics of using fungicides on soybeans.
| |

Fig. 7. Application of fungicides to control Phakopsora pachyrhizi in an efficacy trial located in Paraguay, at the Centro Regional de Investigación Agrícola, one of the research stations for the Ministry of Agriculture and Livestock in Paraguay. |
|
Control of soybean rust can be accomplished through utilization of fungicides and selection away from super susceptible cultivars. Partial resistance or tolerance may be the most effective resistance strategies, especially when combined with stacked single genes or single genes with broad resistance. The scenario that is playing out now in Brazil will provide information needed to develop effective control strategies before the pathogen arrives in the continental U.S. Each season without the introduction of the disease into the U.S. allows for additional fungicides to be evaluated and registered, application methods to be improved, and provides time for the incorporation of resistance and/or tolerance into commercial germplasm.
Acknowledgement and Disclaimer
Photo credits: USDA-Agricultural Research Service except where noted from collaborators in the figure legend.
Names are necessary to report factually on available data; however, the USDA neither guarantees nor warrants the standard of the product, and the use of the name by the USDA implies no approval of the product to the exclusion of others that may also be suitable.
Additional Resources
Pest Alert: Soybean Rust
from PPQ-APHIS-USDA
Diagnostic Methods for Soybean Rust Identification
from PPQ-APHIS-USDA
Soybean Rust
from the Plant Health Initiative
New Soybean disease in South Africa
from the Southern African Society for Plant Pathology (SASPP)
Pest Alert: Soybean Rust
from the North Central Pest Management Center
Literature Cited
1. Anonymous. 1983. Progress Report: Asian Vegetable Research and Development Center.
2. Anonymous. 1992. Breeding for high yield potential combined with soybean rust tolerance. Shanhua, Tainan, Taiwan: Asian Vegetable Research and Development Center.
3. Akinsanmi, O. A., Ladipo, J. L., and Oyekan, P. O. 2001. First report of soybean rust (Phakopsora pachyrhizi) in Nigeria. Plant Dis. 85:97.
4. Bonde, M. R., Melching, J. S., and Bromfield, K. R. 1976. Histology of the suscept-pathogen relationship between Glycine max and Phakopsora pachyrhizi, the cause of soybean rust. Phytopathology 66:1290-1294.
5. Bromfield, K. R. 1980. Soybean rust: some considerations relevant to threat analysis. Prot. Ecol. 2:251-257.
6. Bromfield, K. R. 1984. Soybean rust, Monograph (American Phytopathological Society), No. 11. American Phytopathological Society. St. Paul, MN.
7. Bromfield, K. R., and Hartwig, E. E. 1980. Resistance to soybean rust [Phakopsora pachyrhizi] and mode of inheritance. Crop Sci. 20:254-255.
8. Bromfield, K. R., and Hartwig, E. E. 1980. Resistance to soybean rust and mode of inheritance. Crop Sci. 20:254-255.
9. Bromfield, K. R., Melching, J. S., and Kingsolver, C. H. 1980. Virulence and aggressiveness of Phakopsora pachyrhizi isolates causing soybean rust. Phytopathology 70:17-21.
10. Burdon, J. J., and Marshall, D. R. 1981. Inter- and intra-specific diversity in the disease-response of Glycine species to the leaf-rust fungus Phakopsora pachyrhizi. J. Ecol. 69:381-390.
11. Burdon, J. J., and Speer, S. S. 1984. A set of differential Glycine hosts for the identification of races of Phakopsora pachyrhizi Syd. Euphytica 33:891-896.
12. Desborough, P. J. 1984. Selection of soybean cultivar and sowing date as a strategy for avoidance of rust (Phakopsora pachyrhizi Syd.) losses in coastal New South Wales. Aust. J. Exp. Agric. Anim. Husb. 24:433-439.
13. Frederick, R. D., Snyder, C., Peterson, G. L., and Bonde, M. R. 2002. Polymerase chain reaction assays for the detection and descrimination of the soybean rust pathogens Phakopsora pachyrhizi and P. meibomiae. Phythopathology 92:217-227.
14. Hartman, G. L. 1995. Highlights of soybean rust research at the Asian Vegetable Research and Development Center. Pages 19-28 in: Soybean Rust Workshop, 9-11 August 1995. J. B. Sinclair and G. L. Hartman, eds. College of Agriculture, Consumer, and Environmental Sciences, National Soybean Research Laboratory Publication Number 1, Urbana, Illinois
15. Hartman, G. L., Wang, T. C., and Hymowitz, T. 1992. Sources of resistance to soybean rust in perennial Glycine species. Plant Dis. 76:396-399.
16. Hartman, G. L., Saadaoui, E. M., and Tschanz, A. T., Scientific eds. 1992. Annotated bibliography of soybean rust (Phakopsora pachyrhizi Sydow), AVRDC Library Bibliography Series 4-1, Tropical Vegetable Information Service. Taipei: Asian Vegetable Research and Development Center.
17. Hartwig, E. E. 1986. Identification of a fourth major gene conferring resistance to soybean rust. Crop Sci. 26:1135-1136.
18. Hartwig, E. E., and Bromfield, K. R. 1983. Relationships among three genes conferring specific resistance to rust in soybeans. Crop Sci. 23:237-239.
19. Javaid, I., and Ashraf, M. 1978. Some observations on soybean diseases in Zambia and occurrence of Pyrenochaeta-Glycines on certain varieties. Plant Dis. Rep. 62:46-47.
20. Killgore, E., and Heu, R. 1994. First report of soybean rust in Hawaii. Plant Dis. 78:1216.
21. Koch, E., and Hoppe, H. H. 1988. Development of infection structures by the direct-penetrating soybean rust fungus (Phakopsora pachyrhizi Syd.) on artificial membranes. J. Phytopathol. 122:232-244.
22. Koch, E., Ebrahim Nesbat, F., and Hoppe, H. H. 1983. Light and electron microscopic studies on the development of soybean rust (Phakopsora pachyrhizi Syd.) in susceptible soybean leaves. Phytopathol. Z. 106:302-320.
23. Kochman, J. K. 1977. Soybean rust in Australia. 44-48 in: Rust of Soybean--The problem and research needs. R. E. Ford and J. B. Sinclair, eds. International Agricultural Publications, Manila, the Philippines
24. Kuchler, F., Duffy, M., Shrum, R. D., and Dowler, W. M. 1984. Potential economic consequences of the entry of an exotic fungal pest: the case of soybean rust. Phytopathology 74:916-920.
25. Levy, C., Techagwa, J. S., and Tattersfield, J. R. 2002. The status of soybean rust in Zimbabwe and South Africa. Paper read at Brazilian Soybean Congress, at Fozdo Iguacu, Prarana, Brazil.
26. Lin, S. Y. 1966. Studies on the physiologic races of soybean rust fungus, Phakopsora pachyrhizi Syd. J. Taiwan Agric. Res. 15:24-28.
27. Marchetti, M. A., Uecker, F. A., and Bromfield, K. R. 1975. Uredial development of Phakopsora pachyrhizi in soybeans. Phytopathology 65:822-823.
28. Marchetti, M. A., Melching, J. S., and Bromfield, K. R. 1976. The effects of temperature and dew period on germination and infection by uredo spores of Phakopsora pachyrhizi. Phytopathology 66:461-463.
29. McLean, R., and Byth, D. E. 1976. Resistance of soybean to rust [Phakopsora pachyrhizi] in Australia. Newsl. Aust. Plant Pathol. Soc. 5:34-36.
30. McLean, R., and Byth, D. E. 1980. Inheritance of resistance to rust (Phakopsora pachyrhizi) in soybean. Aust J. Agric. Res. 31:951-956.
31. McLean, R. J. 1979. Histological studies of resistance to soybean rust, Phakopsora pachyrhizi Syd. Aust. J. Agric. Res. 30:77-84.
32. Melching, J. S., Dowler, W. M., Koogle, D. L., and Royer, M. H. 1989. Effects of duration, frequency, and temperature of leaf wetness periods on soybean rust. Plant Dis. 73:117-122.
33. Morel, W., and Yorinori, J. T. 2002. Situacion de la roja de la soja en el Paraguay. Bol de Diulgacion No. 44. Ministerio de Agricultura y Granaderia, Centro Regional de Investigacion Agricola, Capitan Miranda, Paraguay.
34. Ogle, H. J., Byth, D. E., and McLean, R. 1979. Effect of rust (Phakopsora pachyrhizi) on soybean yield and quality in south-eastern Queensland. Aust. J. Agric. Res. 30:883-893.
35. Ogle, H. J., Byth, D. E., and McLean, R. 1979. Effect of rust (Phakopsora pachyrhizi) on soybean yield and quality in Eastern Australia. Soybean Rust Newsletter.
36. Ono, Y., Buritica, P., and Hennen, J. F. 1992. Delimitation of Phakopsora, Physopella and Cerotelium and their species on Leguminosae. Mycol. Res. 96:825-850.
37. Patil, P. V., and Anahosur, K. H. 1998. Control of soybean rust by fungicides. Indian Phytopathology 51:265-268.
38. Poonpolgul, S., and Surin, P. 1985. Physiological races of soybean rust in Thailand. Thai Phytopathology 5:119-120.
39. Rossi, R. L. 2003. First report of Phakospora pachrhizi, the causal organism of soybean rust in the Provence of Misiones, Argentina. Plant Dis. 87:102.
40. Rytter, J. L., Dowler, W. M., and Bromfield, K. R. 1984. Additional alternative hosts of Phakopsora pachyrhizi, causal agent of soybean rust. Plant Dis. 68:818-819.
41. Saksirirat, W., and Hoppe, H. H. 1991. Teliospore germination of soybean rust fungus (Phakopsora pachyrhizi Syd.). J. Phytopathol. 132:339-342.
42. Sharma, N. D., and Mehta, S. K. 1996. Soybean rust in Madhya Pradesh. Acta Botanica Indica 24:115-116.
43. Shin, D. C., and Tschanz, A. T. 1986. Studies on physiological reactions of soybean cultivars tolerant and susceptible to rust (Phakopsora pachyrhizi Syd.). Korean J. Crop Sci. 31:440-446.
44. Sinclair, J. B. 1977. Control of soybean rust [Phakopsora pachyrhizi] by means other than breeding for resistance [Fungicides]. INTSOY Ser Int Soybean Program 12:85-88.
45. Sinclair, J. B., and Hartman, G. L., eds. 1995. Soybean Rust Workshop, 9-11 August 1995, College of Agriculture, Consumer, and Environmental Sciences,National Soybean Research Laboratory Publication Number 1, Urbana, Illinois.
46. Singh, B. B., Gupta, S. C., and Singh, B. D. 1975. Sources of field resistance to rust [Phakopsora pachyrihizi] and yellow mosaic diseases of soybean. Indian J. Genet. and Plant Breed. 34:400-404.
47. Vakili, N. G. 1979. Field survey of endemic leguminous hosts of Phakopsora pachyrhizi in Puerto Rico. Plant Disease Reporter 63:931-935.
48. Vyas, S. C., Rajasekaran, G., and Geetha, M. 1997. Soybean Rust. SOPA Digest 5:33-41.
49. Wang, T. C., and Hartman, G. L. 1992. Epidemiology of soybean rust and breeding for host resistance. Pl. Prot. Bull. 34:109-124.
50. Yang, X. B. 1995. Assessment and management of the risk of soybean rustProceeding of the soybean rust workshop, 9-11 August 1995. J. B. Sinclair and G. L. Hartman, eds. National Soybean Research Laboratory, Urbana, IL
51. Yeh, C. C., Tschanz, A. T., and Sinclair, J. B. 1981. Induced teliospore formation by Phakopsora pachyrhizi on soybeans and other hosts. Phytopathology 71:1111-1112.

