Gergerich, R.C., and V. V. Dolja. 2006. Introduction to Plant Viruses, the Invisible Foe.
The Plant Health Instructor. DOI: 10.1094/PHI-I-2006-0414-01
Rose C. Gergerich1 and Valerian V. Dolja2
1Department of Plant Pathology, University of Arkansas, Fayetteville, AR
2Department of Botany and Plant Pathology, Oregon State University, Corvallis, OR
Introduction
Viruses are infectious pathogens that are too small to be seen with a light microscope, but despite their small size they can cause chaos. The simplest viruses are composed of a small piece of nucleic acid surrounded by a protein coat. As is the case with other organisms, viruses carry genetic information in their nucleic acid which typically specifies three or more proteins. All viruses are obligate parasites that depend on the cellular machinery of their hosts to reproduce. Viruses are not active outside of their hosts, and this has led some people to suggest that they are not alive. All types of living organisms including animals, plants, fungi, and bacteria are hosts for viruses, but most viruses infect only one type of host. Viruses cause many important plant diseases and are responsible for losses in crop yield and quality in all parts of the world.
The purpose of this chapter is to provide an overview of the fascinating microscopic world of plant viruses and to describe the basic concept of a virus, the structure of virus particles and genomes, virus life cycles, the evolution and diversity of plant viruses, as well as the common manifestations of plant virus diseases and major approaches to managing these diseases. We hope to convey to the reader our grudging admiration for these small pathogens and for their success in manipulating their plant hosts so successfully.
History
The beginnings of plant virology date back to the late 19 th century, when Dutch microbiologist Martinus Beijerinck and Russian researcher Dmitrii Iwanowski were investigating the cause of a mysterious disease of tobacco (Scholthof 2001). These researchers independently described an unusual agent that caused mosaic disease in tobacco (Zaitlin 1998). What distinguished this agent from other disease-causing agents was its much smaller size compared to that of other microbes. This agent, later named
Tobacco mosaic virus (TMV), was the first virus to be described. Since then, a large number of diverse viruses have been found in plants, animals, fungi, and bacteria. The current estimate of recognized viruses is approaching 4,000, of which about 1,000 are plant viruses. The main reason that we study plant viruses is the negative impact that viral diseases have on crop production. Historically, viruses are perceived almost exclusively as a health threat to humans, livestock, and crop plants. However, recent progress in understanding virus-host interactions has transformed viruses into important tools of biomedicine and biotechnology. For instance, plant viruses are being used to produce large quantities of proteins of interest in plants (Pogue et al. 2002) and to develop safe and inexpensive vaccines against human and animal viruses (Walmsley and Arntzen 2000).
Basic Biology
Viruses represent not just another group of pathogens, but rather a fundamentally different form of life. Unlike all other living organisms, viruses are non-cellular. In contrast to cells, which multiply by dividing into daughter cells, viruses assemble from pools of their structural components. Mature virus particles are dormant; they come alive and reproduce only inside infected cells. In other words, viruses are obligate parasites that cannot be cultivated using any growth media suitable for bacterial, fungal, plant or animal cell types. All viruses lack protein-synthesizing and energy-producing apparatuses. As a rule, virus particles are immobile outside the infected host; they rely on the aid of other organisms or the environment for their dissemination.
Morphology
There is a simple structural principle that applies to virtually all viruses in their mature form. Virus particles (virions) are composed of two principal parts, the genome that is made of nucleic acid, and a protective shell that is made of protein. In addition, some virus particles are enveloped by an outer membrane containing lipids and proteins (lipoprotein membrane). The protein shells of plant viruses (capsids) are assembled in accord with one of the two fundamental types of symmetry. The first type of virion is helical (roughly elongated). The elongated viruses come in two major variants, rigid rods (Figure 1) and flexuous filaments (Figure 2). In both of these variants, the nucleic acid is highly ordered: it assumes the same helical conformation as the proteinaceous capsid. The second type of virus particle is icosahedral (roughly spherical; Figure 3); the variations of this basic shape include bacilliform virions (Figure 4) and twin virions composed of two joined incomplete icosahedra (Figure 5). In the icosahedral virions, the genomic nucleic acid forms a partially ordered ball inside the proteinaceous capsid. The icosahedral and elongated virions alike can self-assemble in a test tube if the nucleic acid and protein subunits are incubated under proper conditions.
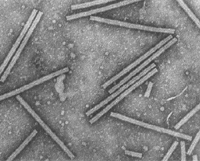
Figure 1 |

Figure 2 |

Figure 3 |
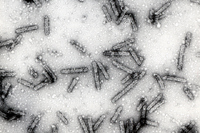
Figure 4 |
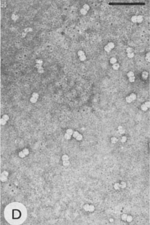
Figure 5 |
Viruses are the smallest among all known organisms. The typical diameter of a spherical plant virus is ~30 nm. The rigid, rod-shaped TMV particle is 300 x 18 nm and consists of an RNA genome of about 6,400 nucleotides encapsidated by 2,130 copies of the TMV coat protein. Some of the filamentous viruses reach the length of ~2000 nm or 2 µm. For comparison, the typical size of a leaf mesophyll cell is ~50 µm.
Reproduction
As in other organisms, the information for virus reproduction is contained within its genome (Figure 6).
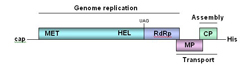
Figure 6 |
Although the genetic material for most organisms is double-stranded (ds) DNA, only a minority of plant viruses possess dsDNA genomes. Some of the plant viruses have genomes that are composed of single-stranded (ss) DNA. However, the majority of plant viruses do not use DNA at all. Instead, the genomes of nearly all plant viruses are made of RNA. Most of these genomes are composed of ssRNA that is the same (positive-sense) polarity as the messenger RNAs of the cell. Some of the RNA viruses use ssRNAs of negative polarity, and yet others have genomes made of dsRNA. Due to this enormous variation in the very nature of the genetic material of viruses, the reproductive cycles and life styles of different viruses are often very distinct from each other.
Since plant viruses are obligate, biotrophic parasites, their life cycles start by penetration of the virion into the cell. Plant viruses are unable to penetrate the plant cuticle and cell wall. It is believed that the virion enters the cytoplasm of the cell passively through wounds caused by mechanical damage to the cuticle and cell wall. The next phase of virus infection is the partial or complete removal of the coat protein shell of the virion in the cytoplasm. Next the cell mediates expression of the viral genome by providing a transcription apparatus (for DNA viruses) and a translation apparatus (for all viruses). The DNA viruses must be transported to the nucleus for transcription in order to gain access to the cell proteins required for the production of messenger RNA from viral DNA. Translation of viral RNA in the cytoplasm produces viral proteins that are required for completion of the virus life cycle.
All viruses must direct the formation of at least three types of proteins: replication proteins that are essential for nucleic acid production, structural proteins that form the protein shell and other components contained in the virions, and movement proteins that mediate virus transport between plant cells (Figure 6). The viral replication proteins combine with cellular proteins to produce a complex of proteins that manufactures multiple copies of the virus genome. These newly made genomes interact with the structural proteins to form new virions. The diagram below (Figure 7) illustrates the steps that occur within the cell during the replication of
Tobacco mosaic virus.
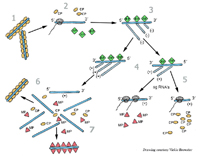
Figure 7 |
The next step in the virus reproduction cycle is movement of the virus into neighboring cells. Depending on the virus, the viral genomes or the virions are transported into neighboring cells through small channels called plasmodesmata that form connections between cells. Many viruses produce movement proteins that modify the plasmodesmata channels and facilitate viral movement into neighboring cells. The following diagram (Figure 8) illustrates the movement of
Tobacco mosaic virus from an infected cell to a neighboring cell.
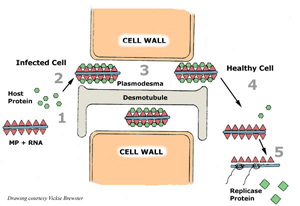
Figure 8 |
The process of cell-to-cell movement is relatively slow: it takes from one to a few hours for a virus to multiply in a cell and move to the next cell. To successfully colonize an entire plant, a virus needs to enter the vascular system of the plant. The process of systemic, or long-distance transport normally proceeds through the phloem sieve elements where viruses move passively with the flow of photosynthates. After quite rapid systemic spread of the virus (centimeters per hour) in the phloem, the virus moves from the phloem into surrounding cells where it reproduces and spreads by cell-to-cell movement. The time between initial infection of one or a few cells and systemic infection of the plant varies from a few days to a few weeks depending on the virus, host plant, and environmental conditions. Transmission of the virus from one plant to another (see section on survival and dissemination) completes the virus life cycle.
Systematics
Ideally, systematics (the study of the kinds of organisms and the relationships between them) should reflect the evolutionary history of a biological species. This history can be constructed by analysis of fossilized predecessors and by comparison of the genes of existing organisms (phylogenetic analysis). Since viruses left no fossils, their evolution can be reconstructed with some confidence only by phylogenetic analysis. This analysis works reasonably well at the intermediate taxonomic levels, such as virus genus and family. However, due to the nature of the viral form of life, it gets more complicated or even unfeasible at the lower (species) level or higher levels. Because the viruses exhibit very high rates of reproduction and mutation, each reproduction cycle results in a number of genetically similar, but not identical variants. Further selection and evolution of slightly different genomes results in a continuum of variants that range from point mutants to virus strains. Unlike cellular organisms, which are believed to originate from a common ancestor, the viruses most likely originated more than once. The diverse origins of different virus groups permit their evolutionary classification only within particular virus types, such as viruses that have positive-sense RNA genomes, or small ssDNA genomes.
When we consider plant virus classification, we should remember that it was the mid 1930s before the first plant virus was purified and characterized. Prior to this time, most plant virologists named a virus based on the host plant in which it was found and the type of symptom that the virus caused in the plant. For example,
Tobacco mosaic virus was first described in tobacco in which it induces a mosaic pattern in the leaves (Figure 9). Virus names are usually shortened to an acronym, for example,
Tobacco mosaic virus is shortened to TMV, and
Tomato spotted wilt virus is shortened to TSWV. The names of many virus genera and families are derived from an important virus within the family. For example, the family name
Bromoviridae is derived from
Brome mosaic virus which is in this family.

Figure 9 |
The largest subdivisions of plant viruses are defined by the chemical form of their genomes (Plant Viruses Online
http://image.fs.uidaho.edu/vide/refs.htm ). By far, the largest number of plant viruses possess single-stranded (ss), positive-sense RNA genomes, and these viruses are called positive-strand RNA viruses. Examples of the most economically important families of the positive-strand RNA viruses are
Bromoviridae, Closteroviridae, Luteoviridae, and
Potyviridae. Relatively few plant viruses, exemplified by the families
Bunyaviridae and
Rhabdoviridae, possess negative-sense RNA genomes.
Reoviridae is the largest family of the double-stranded (ds) RNA viruses. There is only one family of the plant viruses with dsDNA genomes, the
Caulimoviridae family or so-called pararetroviruses, and replication of these viruses involves an RNA intermediate. The true plant DNA viruses are represented by the large and economically important family
Geminiviridae. These viruses possess ssDNA genomes, and they have a dsDNA intermediate in their life cycle (Hull 2002). The evolutionary relationships among the positive-strand, negative-strand, and dsRNA viruses, as well as the pararetroviruses and ssDNA viruses appear to be extremely distant if not absent.
Survival and Dissemination
Only relatively few viruses, such as TMV, rely on long-term (up to decades) survival in the environment and on passive mechanical transmission from plant to plant (Ford and Evans 2003). Most plant viruses are actively transmitted from infected to healthy plants by a living organism called a vector. Plant-feeding arthropods, nematodes and plant-parasitic fungi are the major types of vector organisms for plant viruses (Walkey 1991). Among these, aphids and whiteflies (Figures 10A, B, C) are capable of transmitting the largest number of virus species. Most viruses are actively transmitted to healthy plants in a matter of seconds, hours, or days by vectors. In addition, some viruses are transmitted from infected plants when these plants are vegetatively propagated, for example, through tubers or grafting, or ‘vertically’ through seeds or pollen from infected plants. This type of transmission is particularly important for virus survival during the winter

Figure 10 |
Transmission of viruses by a vector is a very specific process. Each particular virus can be transmitted by only one vector type (e.g., aphid), and not by another (e.g., whitefly). Conversely, each vector species (e.g., green peach aphid
Myzus persicae), can transmit some viruses (e.g.,
Beet yellows virus) but not others (e.g.,
Citrus tristeza virus), despite the fact that these viruses are genetically very similar to each other.
The interaction between a virus and its specific vector that results in virus transmission varies for different virus vectors. The relationships that have developed between viruses and their vectors are complex and of considerable interest to plant virologists because vectors provide the main method of spread for many viruses that cause severe economic losses. For some virus/vector combinations, the interaction between a virus and its vector is very superficial and is the result of attachment to the external surfaces of the vector mouthparts. For example, viruses in the
Potyvirus genus produce a special protein called helper component that “glues” the virions to aphid stylets. In this case, acquisition of the virus from infected plants and inoculation of virus to healthy plants takes from seconds to minutes (Pirone and Blanc 1996). In contrast, viruses in the family
Luteoviridae circulate in their aphid vector moving through the gut wall, into the body cavity, and then into the saliva of the aphid (Figure 11).

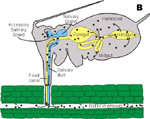
Figure 11 |
It takes at least 12 hours for these viruses to circulate in their aphid vector before they can be transmitted to another plant (Gray and Gildow 2003). For other plant virus vectors, the relationship is very intimate and the viruses actually multiply in the cells of their insect vectors. For example,
Tomato spotted wilt virus multiplies in cells of its thrips vector, and once thrips acquire this virus, they can transmit it for the rest of their life (Sherwood et al. 2003). These plant viruses that multiply in their insect vectors are considered to be both plant and insect viruses. Because management practices for plant diseases are often directed at the virus vectors, an understanding of the virus transmission process is critical for development of effective management strategies for diseases caused by plant viruses
Plant-Virus Interactions
Viruses are capable of infecting virtually all species of cultivated and wild plants. However, host ranges of individual viruses vary from very narrow to very broad. For instance
Citrus tristeza virus infects only a few species in the
Citrus genus, whereas
Cucumber mosaic virus infects over 1000 species in 85 plant families. Susceptibility or resistance of plant species and cultivars to viruses is determined primarily by the plant genotype. Plants possess active and passive means of preventing virus infection. Passive defenses are due to the failure of the plant to produce one or more host factors required for virus reproduction and spread within the host. Active defenses include detection and destruction of the virus-infected cells due to the function of specific resistance genes in the plant. Normally, resistance genes are active only against a particular virus. In addition, plants possess a general defense system that is somewhat analogous to the animal immune system. The major difference between the two is that the immune system in animals targets a pathogen’s proteins, whereas the plant defense system, which is called RNA silencing, detects and degrades viral RNAs (Wassenegger and Pélissier 1998).
Depending on the particular combination of virus and host, and on environmental conditions, a plant’s response to infection may range from a symptomless condition to severe disease and plant death. In some cases, small necrotic or chlorotic spots called local lesions (Figure 12) develop at the site of infection. In most cases, viruses spread throughout the whole plant and cause a systemic infection.

Figure 12 |
Typical leaf symptoms of viral diseases include mosaic patterns (Figure 9), chlorotic or necrotic lesions (Figure 13), yellowing, stripes or streaks (Figure 14, 15), vein clearing, vein banding (Figure 16), and leaf rolling and curling (Figure 17). Flower symptoms include deformation and changes in the color of the flowers including dramatic color mosaics called color breaking (Figure 18, 19). Fruit and vegetable symptoms may include mosaic patterns (Figure 20), stunting, discoloration or malformation (Figure 21), and chlorotic ringspots (Figure 22, 23). Stems of plants may develop stem pitting and grooving or tumors in response to virus infection (Figure 24).

Figure 13 |

Figure 14 |

Figure 15 |

Figure 16 |

Figure 17 |

Figure 18 |

Figure 19 |

Figure 20 |

Figure 21 |

Figure 22 |

Figure 23 |

Figure 24 |
The symptoms induced by plant viruses lead to reduced crop quality and yield. The extent of these crop losses is demonstrated by the following three examples.
Cacao swollen shoot virus (Bowers et al. 2001) is estimated to cause an annual loss of 50,000 tons of cocoa beans in Africa with an estimated value of $28 million dollars. In southeast Asia, infection of rice with
Rice tungro virus leads to an estimated annual economic loss of $1.5 billion dollars annually (Hull 2002).
Tomato spotted wilt virus infects a wide variety of plants including tomato, peanuts, and tobacco (Sherwood et al. 2003), and the estimated annual losses due to infection by this virus worldwide are estimated to be $1 billion dollars (Hull 2002).
The end result of virus infection is a reduction in plant growth, lower yield, inferior product quality, and economic loss to individuals who work in the plant industry. Most of the symptoms induced by viruses can also occur due to adverse environmental conditions or diseases caused by other plant pathogens. Because of this, correct diagnosis of viral diseases normally requires laboratory tests.
Diagnosis of Plant Virus Diseases
Because different viruses may elicit similar symptoms, the disease phenotype can provide only limited, although important, information for disease diagnosis. More specific and reliable methods of virus identification are based on various properties of the virus. These properties and corresponding approaches include the following:
-
Pathogenicity – Bioassays using indicator plants. Some plant genera, such as
Nicotiana (tobacco) and
Chenopodium (lambsquarters) are hosts for a number of viruses. Since the responses of these plants to viral infections under greenhouse conditions are consistent and distinctive, they are commonly used as indicator plants (Walkey 1991). Two major types of responses are local lesions (Figure 12) which are confined to inoculated leaves (local lesion hosts), and systemic infections which produce symptoms on leaves distant from the inoculation site (systemic hosts). Many plant viruses are transmissible to indicator plants by means of mechanical transmission (Figure 25) or grafting.

Figure 25 |
-
Transmissibility – Vector transmission assays. Because of vector specificity, identification of the organism that transmits the virus may provide important information for virus identification.
-
Architecture of virus particles – Electron microscopy. The shape and size of virions distinguish rod-shaped, filamentous, icosahedral, or large enveloped particles (Figures 1-5). On the other hand, viruses sharing the same shape and size are difficult to distinguish by their appearance. For instance, small spherical viruses may be difficult to distinguish from each other and from plant ribosomes.
-
Presence of virus-specific structures in infected cells – Electron microscopy. Because of their intimate association with components of the cell, viruses often form unusual structures within plant cells as a result of virus infection. For example, viruses in the family
Potyviridae produce ‘pinwheel’ inclusions (Figure 26) that are characteristic of viruses in this family and are not found in healthy cells or cells infected with other viruses. Virus-specific inclusions have been characterized for a number of plant virus families and genera, and the detection of these inclusions indicates the presence of a virus within that group.
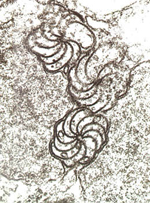
Figure 26 |
-
Properties of the protein coat – Immunological procedures. These tests rely on identification of a virus (the
antigen) through its reaction with specific
antibodies. Specific antibodies are produced by an animal when foreign proteins are introduced into the animal. To obtain antibodies that react specifically with a particular plant virus, scientists inject a purified preparation of the plant virus into an animal (usually a mouse or a rabbit). Several weeks after the injection, blood is collected from the animal and antibodies are separated from the blood. These antibodies react specifically to the viral proteins that were injected into the animal, and scientists have taken advantage of this specific interaction to develop laboratory tests for plant viruses.
One of the most widely used diagnostic tests for plant viruses is an antibody-based procedure called the Enzyme-Linked Immunosorbent Assay (ELISA). An example of one type of ELISA test is illustrated in Figure 27. In this test, an extract of plant tissue being tested for virus is incubated in the well of a plastic microtiter plate, and virions present in the extract bind to the surface of the well. Next, antibody to the virus is added to the well, and the antibody binds to the virus particles. A second antibody that that has been conjugated, or linked, to an enzyme is added to the well where it attaches to the first antibody. Following a wash to remove unbound enzyme-antibody conjugate, a colorless substrate for the enzyme is added. The enzyme processes the substrate and produces a colored product (e.g., yellow). The appearance of color indicates the presence of virus and the intensity of the color can be used to estimate the virus concentration. ELISA is by far the most widely used immunological technique for virus identification in agriculture.
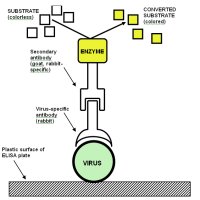
Figure 27 |
-
Properties of viral nucleic acid – PCR amplification. Polymerase Chain Reaction (PCR) is a very sensitive and specific technique for virus detection that is based on the presence of unique nucleic acid sequences in the genome of a virus (Bartlett 2003). The viral DNA sequence that a researcher wants to amplify undergoes about thirty cycles of copying in a small test tube. An example of one type of PCR test is illustrated in Figure 28. In the PCR test, short pieces of DNA (primers) that are complementary to portions of the viral nucleic acid are hybridized (attached) to the viral nucleic acid. A heat-stable polymerase enzyme is then used to synthesize multiple copies of a strand of DNA (cDNA) that is complementary to the portion of the viral nucleic acid sequence between the two primers. During the PCR reaction, the number of molecules of the target sequence doubles with each cycle, and after 30 cycles over 10 billion copies of the target sequence are produced. The PCR product can be detected using gel electrophoresis, a process in which an electrical current is used to separate molecules in a gel based on their size. After separation, the DNA fragments in the gel are stained with a dye that is specific for nucleic acid, and the occurrence of a band of the appropriate size indicates the presence of the virus for which the DNA primers were designed. For RNA viruses, the viral RNA strand is "reverse" transcribed into its DNA complement, followed by amplification of the resulting DNA using PCR
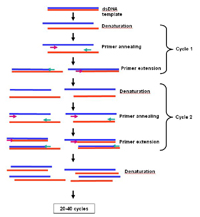
Figure 28 |
Management of Plant Virus Diseases
Although there are virtually no antiviral compounds available to cure plants with viral diseases, efficient control measures can greatly mitigate or prevent disease from occurring. Virus identification is a mandatory first step in the management of a disease caused by a virus. The subsequent strategy for management will depend on the means by which a particular virus enters a crop, how the virus is transmitted between plants within a crop, and how the virus survives when the crop is not being grown (Haddidi et al. 1998). Preventative measures may include use of certified virus-free seed or vegetative stocks, elimination of the virus reservoirs in the surrounding wild vegetation, and modification of planting and harvesting practices. If the virus is known to be transmitted by a particular vector, control or avoidance of this vector is of paramount importance. For instance, insect, nematode or fungal vectors can be controlled by insecticides, nematicides, or fungicides, respectively.
An alternative strategy for virus control is utilization of natural or engineered resistance to virus infection. If available, natural virus resistance genes can be introduced into crop cultivars by conventional breeding. These natural resistance genes are often found among the many cultivars that are available for a particular crop, or the resistance genes are found in related plants that have been identified near the center of origin for that crop plant. Genetic engineering, the transfer of genes between specific organisms using enzymes and laboratory techniques rather than biological hybridization, allows cross-species introduction of such genes. Furthermore, virus resistance can be designed by pre-setting the plant RNA silencing defense system. This is achieved by introduction of fragments of viral nucleic acid into the plant chromosomes. Such transgenic resistance confers immunity to infection by the virus from which the nucleic acid was derived. The effectiveness of this type of transgenic resistance is illustrated by the successful management of
Papaya ringspot virus in Hawaii (Gonsalves et al. 2004). Although genetic engineering offers unlimited opportunities for generating virus-resistant crop plants, its large-scale application has met some resistance from researchers, supervising agencies, and the public. A better understanding of the potential risks associated with introduction of genetically modified plants into the environment is needed to help assess the safety of plants that have been genetically engineered for virus resistance.
Viroids: Infectious Nucleic Acids
Viroids are infectious RNAs that cause a number of economically important plant diseases (Hull 2002). These pathogens are similar to some plant viruses in that they contain an RNA genome, but they differ from RNA plant viruses in two key ways. First, viroids are composed of “naked” RNAs, that is, they lack a protein coat. Secondly, viroids do not produce any proteins when they infect a plant cell despite the fact that they are made of RNA. The RNA genome of viroids is a small, circular molecule that contains between 246 and 375 nucleotides. Even though viroids do not produce their own proteins, they are capable of using the host cell machinery to reproduce their RNA and move into other cells to infect the whole plant.
For many years prior to the discovery that viroids are unique pathogens and distinct from plant viruses, the diseases caused by viroids were classified as plant virus diseases. One reason for this confusion is that the types of symptoms induced by viroids in plants are similar to those induced by plant viruses. These symptoms can be visualized by listing the imaginative names given to some viroids (note that viroids are named in a manner similar to plant viruses, but that the name ends in “d”):
Potato spindle tuber viroid (PSTVd),
Apple scar skin viroid (ASSVd) (Figure 29),
Avocado sunblotch viroid (ASBVd), and
Pear blister canker viroid (PBCVd). Despite their small size, the economic effects of viroids can be devastating. It is estimated that more than 30 million coconut palms in the Philippines have died of the disease caused by
Coconut cadang-cadang viroid (CCCVd) (Figure 30 ).

Figure 29 |

Figure 30 |
Viroids can be classified into two major families, the
Pospiviroidae and the
Avsunviroidae, based on where the viroid replicates in the cell (Tabler and Tsagris 2004). Viroids in the
Pospiviroidae family replicate in the nucleus while viroids in the
Avsunviroidae family replicate in the chloroplast. Because viroids don’t produce any of their own proteins, they depend on one of the plant’s RNA polymerases and other plant proteins for their replication.
Diagnostic tests used to detect viroids do not rely on immunological assays because viroids do not produce any proteins during infection. The diagnostic procedures for viroids are based on bioassays, such as mechanical transmission to indicator plants that produce diagnostic symptoms, and detection procedures based on the viroid RNA. These nucleic acid based tests include gel electrophoresis to detect the viroid RNA, nucleic acid hybridization tests and polymerase chain reaction (PCR) assays.
Viroids are spread to other plants through vegetative propagation, mechanical contamination, pollen and seed. Management strategies for viroid diseases are similar to strategies used for plant viruses, and these include the use of viroid-tested propagation material, use of viroid-free seed, prevention of mechanical contamination, and the use of plants that are resistant to viroid infection.
Conclusions
Plant viruses and viroids are diverse and unusual groups of plant pathogens that infect and cause disease in many crop plants. Because these pathogens depend on the normal cellular machinery of their plant host for reproduction, it is difficult to eliminate them without damaging the host plant. Therefore, most management strategies for diseases caused by plant viruses and viroids are directed at preventing infection of the plant.
References
- Bartlett, J. 2003. PCR Protocols. Humana Press. Totowa, NJ.
- Bowers, J.H., B.A.Bailey, P.K. Hebbar, S. Sanogo, and R.D. Lumsden. 2001. The impact of plant diseases on world chocolate production. Online. Plant Health Progress DOI:10.1094/PHP-2001-0709-01-RV.
- D'Arcy, C.J. and L.L. Domier. 2000. Barley yellow dwarf. The Plant Health Instructor. DOI:10.1094/PHI-I-2000-11-03.
http://www.apsnet.org/edcenter/intropp/lessons/viruses/Pages/BarleyYelDwarf.aspx
- Ford, R. and T. Evans. 2003.
Tobacco mosaic virus. The Plant Health Instructor. DOI: 10.1094/PHI-K-2003-0528-01.
http://www.apsnet.org/edcenter/K-12/TeachersGuide/TobaccoMosaicVirus/Pages/default.aspx
- Gonsalves, D., C. Gonsalves, S. Ferreira, K. Pitz, M. Fitch, R. Manshardt, and J. Slighton. 2004. Transgenic virus resistant papaya: from hope to reality for controlling
Papaya ringspot virus in Hawaii.
http://www.apsnet.org/publications/apsnetfeatures/Pages/papayaringspot.aspx
- Gray, S., and Gildow, F. 2003. Luteovirus-aphid interactions. Annual Review of Phytopathology 41: 539-566.
- Haddidi, A., R.K. Khertarpal, and H. Koganezawa, (eds.). 1998. Plant Virus Disease Control. APS Press, St. Paul, MN.
- Hull, R. 2002. Matthews' Plant Virology. 4th Edition. Academic Press, San Diego, CA.
- Levy, L., V. Damsteegt, R. Scorza, and M. Kolber. 2000. Plum pox potyvirus disease of stone fruits.
http://www.apsnet.org/edcenter/intropp/PathogenGroups/Pages/PlantViruses.aspx
- Pirone, T.P. and S. Blanc. 1996. Helper-dependent vector transmission of plant viruses. Annual Review of Phytopathology 34: 227-247.
- Pogue, G.P., J.A. Lindbo, S.J. Garger, and W.P. Fitzmaurice. 2002. Making an ally from an enemy: plant virology and the new agriculture. Annual Review of Phytopathology 40: 45-74.
- Scholthof, K-B.G. 2000. Tobacco mosaic. The Plant Health Instructor. DOI:10.1094/PHI-I-2000-1010-01.
http://www.apsnet.org/edcenter/intropp/lessons/viruses/Pages/TobaccoMosaic.aspx
- Scholthof, K.-B. G. 2001. 1898 – the beginning of virology…time marches on. The Plant Health Instructor. DOI:10.1094/PHI-I-2001-0129-01.
http://www.apsnet.org/edcenter/intropp/lessons/viruses/Pages/TobaccoMosaic.aspx
- Sherwood, J.L., T.L. German, J.W. Moyer, and D.E. Ullman. 2003. Tomato spotted wilt. The Plant Health Instructor. DOI:10.1094/PHI-I-2003-0613-02.
http://www.apsnet.org/edcenter/intropp/lessons/viruses/Pages/TomatoSpottedWilt.aspx
Tabler, M. and M. Tsagris. 2004. Viroids: petite RNA pathogens with distinguished talents. Trends in Plant Science 9: 339-348. - Walkey, D. 1991. Applied Plant Virology. 2nd edition. Chapman and Hall, London.
- Walmsley, A.M. and C.J. Arntzen. 2000. Plants for delivery of edible vaccines. Current Opinion in Biotechnology 11: 126-129.
- Wassenegger, M., and T. Pélissier. 1998. A model for RNA-mediated gene silencing in higher plants. Plant Molecular Biology 37: 349-362.
- Wintermantel, W.M. 2004. Emergence of greenhouse whitefly (Trialeurodes vaporariorum) transmitted Criniviruses as threats to vegetable and fruit production in North America.
http://www.apsnet.org/publications/apsnetfeatures/Pages/GreenhouseWhitefly.aspx
- Zaitlin, M. 1998. The discovery of the causal agent of tobacco mosaic disease. Pages 105-110 in: Discoveries in Plant Biology. S.D. Kung and S.F. Yang, eds. World Publishing Co. Ltd. Hong Kong.

