Fletcher, J. and A. Wayadande. 2002. Bacterias Fastidiosas Colonizadoras Vasculares. Trans. José Carlos Ureta R. 2009. The Plant Health Instructor. DOI: 10.1094/PHI-I-2009-0323-01
Jacqueline Fletcher and Astri Wayadande
Department of Entomology and Plant Pathology, Oklahoma State University, Stillwater, OK
Traductor: José Carlos Ureta R.
Departamento de Protección Vegetal – Facultad de Ciencias Agropecuarias, Universidad de Panamá, Panamá.
Traducción revisada por Ana María Romero. Fitopatología-Facultad de Agronomía, Universidad de Buenos Aires, Argentina.
La mayoría de las bacterias fitopatogénicas invaden a su planta hospedera a través de aberturas naturales o heridas, colonizando los espacios intercelulares, expresando diferentes factores de patogenicidad e induciendo diversas reacciones en la planta afectada. Sin embargo, unas pocas son introducidas directamente dentro de los tubos cribosos ricos en azúcares del floema o en los haces que transportan agua y nutrientes del xilema por insectos que se alimentan de dichos tejidos vasculares. Estas bacterias utilizan estrategias inusuales y algunas veces únicas, por las cuales optimizan su ocupación en dichos nichos y así obtienen su alimentación por parte de la planta afectada. Su localización dentro de tejidos vivos (vasos cribosos) o tejidos degenerados (elementos del xilema) de la planta, en vez de los espacios intercelulares, hace que se les presenten retos y oportunidades para evitar los mecanismos de defensa de la planta. Además, muchas de las bacterias que afectan al sistema vascular de las plantas establecen una estrecha relación con estos tejidos. Estas relaciones pueden ser benéficas o perjudiciales en relación a su insecto vector. Estos patógenos inusuales y sus múltiples hospedantes proveen ejemplos fascinantes de las complejas redes de interacción entre organismos. Las bacterias colonizadoras de tejidos vasculares pueden agruparse en tres grupos: a.- Molicutes sin pared celular (fitoplasmas y espiroplasmas), b.- Bacterias con pared celular habitantes del floema, c.- Bacterias con pared celular limitadas al xilema.
Molicutes: Fitoplasmas y Espiroplasmas
Historia
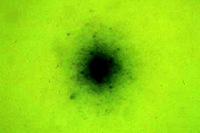
Las plantas con amarillamientos (“yellows”) se caracterizan por presentar un follaje clorótico y falta de crecimiento vigoroso generalizado (Figura 1). También pueden ocurrir otros síntomas como virescencia y asimetría floral, necrosis vascular, proliferación de retoños y yemas, enanismo y muerte. Durante mucho tiempo se creyó que los agentes causales de estas enfermedades eran virus ya que la sintomatología era similar a la causada por virus fitopatógenos, ningún microorganismo podía ser aislado de manera consistente, estos agentes causales eran capaces de pasar a través de filtros que retenían bacterias y no se podía reconocer a ningún microorganismo en observaciones con el microscopio compuesto o de luz. Finalmente, al igual que en el caso de algunas virosis, estos agentes causales eran transmitidos mediante injerto y/o insectos vectores. No fue sino hasta la década de 1960 que algunos “amarillamientos” fueron atribuidos correctamente a fitopatógenos procariotas.
 |
| Figura 1. |
El mayor crédito por un trabajo sobresaliente en lo referente a enfermedades denominadas amarillamientos se debe a L.O. Kunkel, un fitopatólogo cuyas investigaciones dedicadas y meticulosas en amarillamientos del aster (“aster yellows”), se extendieron desde mediados de la década de 1920 hasta mediados de la década de 1950. Kunkel comprobó que el agente causal del amarillamiento del aster es infeccioso y transmitido, en la naturaleza, por chicharritas (“leafhoppers”; Hemiptera: Cicadellidae) que se alimentaban del floema. Familiarizado con insectos transmisores de virus, Kunkel fue capaz de determinar los períodos mínimos de adquisición, incubación e inoculación para el patógeno del amarillamiento del aster en su vector. En una serie de experimentos ingeniosos, mostró que tratando con calor a los vectores infectivos a una temperatura subletal, destruía su capacidad para transmitir el patógeno. Sin embargo, cuando estos insectos tratados e incapaces de transmitir al patógeno se les permitía un periodo de incubación nuevo, la habilidad para transmitirlo se restauraba. Sus datos comprobaron que el patógeno se propagaba dentro del insecto vector. Por sus muchas contribuciones L.O. Kunkel ha sido nombrado como el "Padre de la Micoplasmología en Plantas e Insectos."
No obstante, el entendimiento de que no todos los agentes causales de amarillamientos en plantas eran virus, tuvo que esperar a los famosos experimentos realizados en Japón por fitopatólogos que estudiaron la devastadora enfermedad denominada enanismo de la mora (“mulberry dwarf disease”). A finales de la década de 1960, Doi, deseando visualizar al “virus” que causaba la enfermedad del enanismo de la mora, estudió detenidamente secciones de tejidos de plantas afectadas con la ayuda de un microscopio electrónico, pero no pudo encontrar partículas virales. Por azar, una sesión en el revelador de impresiones fue interrumpida por un microbiólogo veterinario que estaba revelando fotografías del patógeno animal Mycoplasma gallisepticum. Fue Koshimizu quien observó las similitudes entre las fotografías de su micoplasma y los extraños cuerpos del tamaño de mitocondrias en la fotografías de Doi. Aunque reticente, Doi tomó en consideración la sugerencia de Koshimizu para observar si el antibiótico tetraciclina podía afectar el desarrollo de la enfermedad. Cuando el uso de dicho antibiótico conllevó a la remoción de los síntomas, había nacido un campo nuevo en la fitopatología. Aunque inicialmente se los llamó “Pleuropneumonia-like organisms (PPLOs) o Mycoplasma-like organisms (MLOs) debido a su similitud superficial con dichos grupos, desde entonces estos fitopatógenos han mostrado diferencias significativas con los micoplasmas y se los ha designado fitoplasmas (Sears and Kirkpatrick 1994). Luego de su descubrimiento, muchas otras enfermedades que se asumían tener una etiología viral, pero a las que no se les encontraba el virus causal, fueron reexaminadas y en los años subsiguientes el número de fitoplasmas conocidos y enfermedades inducidas por fitoplasmas ha continuado creciendo. Aunque la caracterización de estos organismos se ha visto limitada por la imposibilidad de cultivarlos in vitro, la aplicación de tecnología molecular reciente ha dado como resultado una expansión rápida de nuestro conocimiento acerca de los fitoplasmas y en el establecimiento de un género “candidato” para los fitoplasmas, Candidatus Phytoplasma.
Alrededor de 1970, la investigación acerca del achaparramiento ó atrofiamiento del maíz (“corn stunt”), una enfermedad significativa en los trópicos y subtrópicos, fue lo que impulsó el descubrimiento de otro grupo de fitopatógenos relacionados. Davis y Worley en el Departamento de Agricultura de los Estados Unidos (USDA-ARS en Beltsville, Maryland) estaban tratando de identificar el agente causal del achaparramiento del maíz. Cuando examinaron filtrados de la planta con el microscopio compuesto, observaron células helicoidales pequeñas que no poseían pared celular (Figura 2). Posteriormente llamaron a estos nuevos microorganismos espiroplasmas. A diferencia de los fitoplasmas, los espiroplasmas podían ser cultivados en medios artificiales, lo que facilitó su caracterización en comparación con la de los fitoplasmas.
Biología
Los fitoplasmas, espiroplasmas, micoplasmas y sus organismos relacionados, todos procariotas que carecen de pared celular y que comparten otras características citológicas y moleculares inusuales, forman la clase Molicutes ("molli" - suave; "cute" - piel). Aunque superficialmente similares a las bacterias con pared celular “en forma de L”, las cuales han perdido parte o toda su pared celular, los molicutes son taxonómicamente distintos. En un inicio, los investigadores propusieron que los molicutes eran microbios primitivos, posiblemente los descendientes de bacterias que existieron antes del desarrollo de una pared celular de peptidoglicano. Sin embargo, análisis moleculares posteriores mostraron claramente que los molicutes provenían de antecesores bacterianos Gram positivos con pared celular. El grupo comprende a los organismos celulares autoreplicantes más pequeños y simples, con un diámetro de entre 0.3 y 1.2 µm, y genomas correspondientemente pequeños (108 -109 daltons).
Los molicutes comparten varias características únicas. Debido a que no presentan una cobertura rígida y están rodeados solamente por la membrana plasmática son pleomórficos, su morfología está influenciada por el medio que los rodea, son osmóticamente frágiles y resistentes a antibióticos como la penicilina, la cual inhibe la formación de la pared celular. Los micoplasmas y espiroplasmas, pero no los fitoplasmas, son cultivables en medios artificiales, pero la dificultad para ser cultivados y el requerimiento de esteroles y otros ingredientes poco usuales en el medio de cultivo de muchos de ellos ha llevado a su descripción como “fastidiosos”.
El hecho de que los molicutes posean los genomas conocidos más pequeños ha hecho de este grupo un objetivo para proyectos genómicos. El récord lo posee actualmente un fitoplasma fitopatógeno del Pasto Bermuda con 530 kb; en contraste, el genoma de Escherichia coli es de alrededor de 4600 kb. El genoma de Mycoplasma genitalium (580 kb) fue el primer genoma microbiano completamente secuenciado. Muchos otros genomas de micoplasmas se encuentran disponibles, el de Spiroplasma kunkelii está cercano a su finalización (http://www.genome.ou.edu/spiro.html), y los de otros micoplasmas, espiroplasmas y fitoplasmas se encuentran en curso. Los aspectos moleculares de la biología de los molicutes han sido recientemente revisados por Razin et al. (1998). El genoma de estos procariotas presenta un contenido bajo de G+C (23% - 41%). Los espiroplasmas y micoplasmas, pero no los fitoplasmas, utilizan el codón AUG para codificar el aminoácido triptófano en vez de que sea utilizado como un codón de terminación de la traducción como es utilizado en la mayoría de los seres vivos. El genoma ha tenido un número de supresiones durante su reducción de tamaño, algunas de las cuales sin duda están relacionadas a adaptaciones a sus hábitos parasíticos y patogénicos en los cuales ciertas funciones ya no son requeridas. Sin embargo, muchos molicutes tienen información genética adicional como secuencias de ADN extracromosómico (virus y plásmidos) (figura 3), las cuales aparentemente se integran de manera libre en el genoma y producen diferentes tipos de re-arreglos genómicos.
Los Espiroplasmas (Familia Spiroplasmataceae) usualmente son de morfología helicoidal, miden de 0.08 - 0.2 µm de ancho y 2-4 µm de largo. Las especies fitopatógenas pueden perder su forma helicoidal cuando están dentro de su insecto vector. Su motilidad incluye movimientos translacionales (incluidos aquellos basados en gradientes químicos, temperaturas y otros factores ambientales), rotación sobre un eje central y un movimiento de flexión y volcado.
Desde el descubrimiento de S. kunkelii en plantas de maíz que mostraban síntomas de “achaparramiento” (Figura 4) en los años de la década de 1970, se ha involucrado a los espiroplasmas en otras enfermedades importantes de las plantas. El agente causal de la enfermedad “persistente” ("stubborn") en cítricos, en la cual los árboles afectados desarrollan un hábito de crecimiento disminuido, reducción del tamaño de las hojas y frutos amargos y asimétricos, fue identificado como un espiroplasma y designado con el nombre de Spiroplasma citri. Esta misma especie fue posteriormente aislada de plantas de rábano picante (“horseradish”) en Illinois que presentaban la enfermedad de raíces quebradizas ("brittleroot") (Figura 5), caracterizada por una necrosis del floema, atrofiamiento, y fragilidad de las raíces de la cual se deriva su nombre común. El tercer fitoplasma fitopatógeno conocido es Spiroplasma phoeniceum, el cual fue aislado de plantas de vinca o Hierba Doncella (“periwinkle”; Vinca minor) sembradas como cultivo trampa para S. citri, alrededor de los árboles de cítricos en Siria. El espiroplasma aislado de estas plantas era diferente de S. citri, pero se desconoce su planta (s) hospedante (es) y vector (es) naturales.
 |
 |
| Figura 4. |
Figura 5. |
Aunque los primeros espiroplasmas descubiertos eran fitopatógenos, especies no patogénicas fueron identificadas rápidamente, ocupando nichos como epifitas o causando diversas enfermedades en animales, incluyendo al ser humano. Sin embargo, la gran mayoría vive dentro de insectos. El hecho de que los mosquitos, garrapatas, abejas, Drosophila, escarabajos, chicharritas (“leafhoppers”; Hemiptera: Cicadellidae), moscas y muchas otras especies de insectos porten espiroplasmas, llevó a Hackett y Clark (1989) a sugerir que estos organismos pueden ser uno de los más prevalentes en la tierra. Debido a que estos microbios a menudo causan efectos negativos en el insecto, pueden ser potencialmente útiles para el biocontrol.
Interacción Hospedero-Patógeno
Los espiroplasmas y fitoplasmas causan alrededor de 600 enfermedades en varios cientos de especies de plantas (Calavan y Oldfield 1979). Los amarillamientos caen primordialmente en dos categorías de síntomas. En la primera de ellas, causada por espiroplasmas y algunos fitoplasmas, la sintomatología resulta primariamente de la disfunción del floema y posiblemente también de la competencia entre el patógeno y la planta por los productos elaborados a nivel floemático. Los síntomas incluyen marchitamiento, acortamiento de entrenudos, clorosis foliar, muerte regresiva (“die-back”), amarillamiento o enrojecimiento ocasional del follaje, necrosis del floema y colapso de los vasos cribosos y células acompañantes, deposición de callosa en los elementos del floema, declinación de la planta y muerte. Un segundo tipo, causado por otro grupo de fitoplasmas, involucra desbalances de fitoreguladores de crecimiento que dan como resultado deformaciones muy distintivas y sobrecrecimientos. Esto último incluye el desarrollo temprano y proliferación de retoños axilares (escoba de bruja), crecimientos arbustivos meristemáticos e hinchazones de las nervaduras (Figura 6). Las flores pueden estar parcial o totalmente estériles, y pueden exhibir filodia (pétalos presentando una apariencia de hojas), asimetría, virescencia (pétalos verdes) o quebrado (distribución irregular de los pigmentos en los pétalos) (Figura 7). En un caso, la pigmentación variegada de las brácteas de las flores de Pascua (Poinsettia) se considera una cualidad positiva. Los tipos de síntomas y severidad variarán de acuerdo al patógeno, condiciones ambientales y la edad de la planta en el momento de la infección. Los desastres económicos pueden deberse a una reducción en la calidad del producto o pérdida parcial o total en el rendimiento.
 |
 |
| Figura 6. |
Figura 7. |
Para un manejo efectivo de las enfermedades en las plantas, es crítico un diagnóstico correcto, la detección y la identificación del patógeno. La sintomatología puede ser una herramienta efectiva para un diagnóstico preliminar para algunas enfermedades causadas por molicutes, particularmente por aquellas caracterizadas por irregularidades significativas en su crecimiento, como lo son atrofiamientos, deformación de órganos y cambios en el color de los pétalos. Una decoloración del floema y/o necrosis (Figura 8) también puede indicar la presencia de un patógeno vascular. Sin embargo, se requieren de otros mecanismos más definitivos de identificación para confirmar la etiología de la enfermedad. Cualquiera de los tres espiroplasmas fitopatógenos más conocidos (S. citri, S. kunkelii y S. phoenecium) pueden ser cultivados en medios de cultivo especiales (Figura 9), pero no es así para el caso de los fitoplasmas. En las décadas de 1970 y 1980 se establecieron asociaciones convincentes de fitoplasmas con muchas enfermedades, mediante estudios con el microscopio electrónico, en el cual estos patógenos se observaron en gran número en vasos cribosos de plantas asintomáticas, pero no sanas. Se empleó microcopia de luz en secciones de tejidos de plantas afectadas en combinación con la Tinción de Dienes, la cual tiñe los vasos cribosos colonizados por molicutes (Figura 10), y se adaptó la microscopia de fluorescencia para observar los tejidos teñidos con la tinción para ácido nucleico, DAPI. Más recientemente se ha dispuesto de antisueros para un cierto número de molicutes fitopatogénicos, así que se ha utilizado la técnica de ELISA y otras técnicas inmunológicas. Actualmente, se usan marcadores moleculares (ADN - sonda e imprimador) de diferente sensibilidad y especificidad en la mayoría de los estudios, y en el caso de los fitoplasmas, son complementados con el análisis de perfiles de RFLP para ubicarlos filogenéticamente y establer la relación del agente causal de la enfermedad con molicutes conocidos.
 |
 |
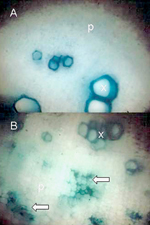 |
| Figura 8. |
Figura 9. |
Figura 10. |
No se entiende a ciencia cierta las interacciones entre los molicutes y las plantas afectadas y la fisiología involucrada que da como resultado el desarrollo de los síntomas observados. Aunque los molicutes evolucionaron de ancestros bacterianos con pared celular, los genes para los mecanismos de patogenicidad encontrados en dichas bacterias pueden o no estar presentes en el reducido genoma de los molicutes. Diferentes factores pueden estar operando en los diferentes sistemas de interacción entre plantas y molicutes, como se evidencia por las dos categorías sintomatológicas mencionadas anteriormente. Ya que los sitios de colonización en el hospedante son los vasos cribosos del floema, es lógico suponer que la interrupción de la función del floema puede ser un factor en la producción de síntomas y la falta de crecimiento vigoroso de la planta afectada.
Sin embargo, ya que los molicutes fitopatogénicos a menudo causan enfermedades en sus respectivos vectores, y de hecho pudieron haberse originado como patógenos de insectos que invadieron un nuevo nicho cuando fueron introducidos por los insectos en el floema de las plantas, es posible que el mecanismo que determina la enfermedad en el insecto opere de la misma manera en la planta afectada.
Posibles factores de virulencia:
- Disfunción del floema. Los espiroplasmas y fitoplasmas se multiplican en poblaciones densas dentro de los vasos cribosos del floema, indudablemente utilizando una gran fracción de los productos de la fotosíntesis elaborados por la planta afectada y que son transportados desde allí a los sitios de crecimiento de la planta. Este tipo de competencia sustrae su cuota en el nivel de crecimiento, altura y producción de flores y frutos. Además, aunque los molicutes pueden y en efecto migran con el flujo de la savia del floema a través de los vasos cribosos, lo más probable es que causen un bloqueo físico del floema, interrumpiendo o inhibiendo la difusión de los nutrientes elaborados. Los depósitos callosos dentro de los vasos cribosos colonizados también pueden contribuir con el bloqueo de estos conductos. Por ejemplo, la savia del floema de las palmas de cocotero que presentaban afecciones por el fitoplasma agente causal del “amarillamiento letal del cocotero”, era más concentrada pero tenía una presión osmótica más baja al compararla con palmas sanas.
- Toxinas. Existe cierta evidencia de que los espiroplasmas producen una sustancia fitotóxica que se transloca y que puede contribuir en la degeneración de las células del floema colonizadas. Una sustancia de bajo peso molecular (< 300), polar, acídica, causó una necrosis en un ensayo con hojas de fríjol, pero su inestabilidad in vitro ha impedido su caracterización detallada.
- Hormonas y reguladores de crecimiento. Los molicutes inducen deformaciones en las hojas, tallos y flores, floración prematura y/o indeterminada, filodia, virescencia y fasciación lo que sugiere que reguladores de crecimiento, producidos ya sea por el patógeno o por la planta, se encuentran desbalanceados. En un experimento se cultivó el meristema apical de una planta infectada con el fitoplasma del amarillamiento del aster o con S. citri, y se determinó que la infección altera los niveles normales de citoquinina y auxina endógenas. La floración prematura causada por el fitopatógeno de la virescencia en la vinca o Hierba Doncella (“periwinkle”), cuyo vector es la chicharrita de la remolacha, puede ser simulado con aplicaciones exógenas de quinetina.
- Enzimas. Normalmente en las enfermedades producidas por molicutes no se presentan síntomas de pudriciones blandas y no existe evidencia experimental en cuanto a la producción de celulasas o pectinasas por parte de fitoplasmas y espiroplasmas. Otras enzimas, sin embargo, pueden intervenir en el desarrollo de la enfermedad. Se han encontrado diferencias en los niveles de esterasas, las cuales desdoblan la suberina, y ribonucleasas en razas patógenas y no patógenas de S. citri.
- Procesos metabólicos. Los síntomas tipo amarillamientos pueden reflejar cambios fisiológicos y efectos en los componentes o en los reguladores de mecanismos metabólicos. Mutantes obtenidos con transposones aislados de S. citri, fallaron en causar síntomas en plantas experimentales y tuvieron una tasa de multiplicación reducida in planta. El transposón fue integrado en un sólo sitio del operón fructosa, en un gen que codifica una proteína que tiene homología con un represor del operón desoxirribonucleótido de E. coli, sugiriendo que el metabolismo de la fructosa en alguna forma estaría relacionado con el proceso infeccioso de este espiroplasma.
- Plásmidos. En casi todas las especies de espiroplasmas en donde se ha investigado se ha encontrado la presencia de plásmidos. Los plásmidos de bacterias patógenas a menudo contienen genes relacionados con su virulencia; sin embargo, la función de estos elementos de ADN extracromosómicos en los molicutes, los cuales presentan un rango de tamaño de 2 a 39 kb (Razin et al.. 1998), es mayormente desconocida. Un plásmido de un espiroplasma ha sido asociado con su resistencia al antibiótico eritromicina.
- Virosis. Los virus en espiroplasmas, cuya morfología varía desde formas bacilares hasta forma de fagos, son prevalentes en espiroplasmas y pueden desempeñar un papel de manera directa o indirecta en la adaptación de los espiroplasmas a sus hospedantes (Razin 1998). Directamente, por supuesto, los virus se pueden replicar y causar lisis en las células de su hospedante (espiroplasma), formando placas en los cultivos bacterianos. El estado del virus (lítico o lisogénico) puede influir en la capacidad del espiroplasma para colonizar, desarrollarse y causar una enfermedad en la planta o el insecto vector, y se ha considerado en cierta medida el uso de estos virus para el control biológico de estos patógenos. Un papel indirecto de los virus en la adaptación de los espiroplasmas surge de la tendencia del ácido nucleico viral a integrarse al azar pero frecuentemente, tanto como un genoma viral completo o como fragmentos, en el genoma del espiroplasma. Estas inserciones pueden inactivar genes o reguladores de genes alterando la biología del espiroplasma, y también proveen un mecanismo para los re-arreglos y supresiones frecuentes y significativas que conllevan a una inesperada alta tasa de cambios evolutivos y adaptaciones a diferentes nichos que presentan estos procariotas sin pared celular.
- Respuesta del Hospedante. Plantas de Hierba Doncella inoculadas con S. citri, Candidatus Phytoplasma aurantifolia, y el fitoplasma del stolbur expresaron diferencialmente por lo menos 24 cADNs de genes relacionados con la fotosíntesis, translocación de azúcares, respuestas a estreses y síntesis de esteroles. Se efectuaron las correlaciones entre algunos cADNs y ciertos síntomas de la enfermedad, aunque las expresiones no fueron únicas para un molicute en particular.
Diseminación
Los molicutes fitopatogénicos son transmitidos por chicharritas del tipo cicadélidos ( “leafhoppers”; Hemiptera, Cicadellidae) y fulgóridos (“planthoppers”; Hemiptera, Fulgoroidea) o por psílidos (Hemiptera, Psyllidae). Estos pequeños insectos se alimentación introduciendo su aparato bucal dentro de los tejidos del mesófilo, xilema o floema de una determinada planta hospedante. La penetración en el floema les permite ingerir la savia del floema por varias horas, periodo durante el cual pueden adquirir espiroplasmas o fitoplasmas de plantas infectadas. Diversos factores pueden influir en la capacidad de uno de estos insectos para que funcione como un vector de dichos fitopatógenos, incluyendo el ser propensos a alimentarse de determinada planta hospedante, su capacidad para mantener la ingestión de la savia del floema y adecuada capacidad para que el espiroplasma o fitoplasma se multiplique en ellos.
Los espiroplasmas y fitoplasmas presentan una relación propagativa con sus insectos vectores (Fletcher et al.. 1998). Luego de ser ingeridos por los insectos vectores, durante la ingestión de la savia del floema, los espiroplasmas o fitoplasmas se ubican en la cavidad corporal del insecto, por la región media del canal alimenticio del intestino. Se piensa que se ubican dentro de las células del insecto mediante un proceso de internalización mediado por un receptor, como se muestra el caso de S. citri (Figura 11). Una vez dentro de las células del epitelio del canal alimenticio, los molicutes se multiplican y posteriormente se movilizan a través del plasmolema y lámina basal de las células del insecto hacia su sistema circulatorio. Utilizando a la hemolinfa como medio de transporte y multiplicación, los mencionados patógenos migran hacia las glándulas salivares de donde son expulsados junto con la saliva hacia el floema en el subsiguiente periodo alimenticio del insecto. El proceso completo de transmisión puede tomar de 2 - 4 semanas, dependiendo la concentración de la fuente y la duración del periodo de adquisición en la planta afectada. Los molicutes fitopatógenos no se les conoce ser transmitidos a la progenie (transmisión transovarial).
 |
| Figura 11. |
S. kunkelii es transmitido en la naturaleza por un grupo específico de cicadélidos del maíz del género Dalbulus. El principal vector en el campo de S. kunkelii es D. maidis (Figura 12), la chicharrita del maíz. El achaparramiento del maíz puede ser devastador para las regiones centrales de México donde cultivan maíz, y debido a los hábitos migratorios de D. maidis, ocasionalmente en las regiones del sur de los Estados Unidos. Otros vectores de esta enfermedad son D. elimatus, un cicadélido restringido a las elevaciones más altas del centro de México, y Macrosteles quadrilineatus, un vector experimental importante, aunque no es un vector natural. La eficiencia en la transmisión puede ser muy alta; 90% - 100% de los individuos de D. maidis que adquieren el patógeno pueden transmitirlo.
S. citri ocurre en todas las regiones productoras de cítricos importantes del mundo. En los Estados Unidos, el principal vector en el campo es la chicharrita de la remolacha, Circulifer tenellus, mientras que en Europa, Circulifer congeners and Scaphitopius spp. son sus vectores más importantes. A diferencia del caso del vector de S. kunkelii, D. maidis, que sólo se alimenta de maíz, C. tenellus presenta un amplio rango de hospederos y puede alimentarse de diferentes grupos de plantas, lo que parcialmente explica el por que S. citri tiene un número de plantas hospedantes de diferentes familias botánicas.
Los fitoplasmas son transmitidos de la misma manera que los espiroplasma, pero sus vectores no se limitan a los cicadélidos. Algunos fitoplasmas, incluyendo a los agentes causales del amarillamiento letal de cocotero y la declinación del peral son transmitidos por fulgóridos y psílidos, respectivamente. Los cicadélidos transmiten a los fitoplasmas eficientemente; por ejemplo, D. maidis, el vector del fitoplasma del atrofiamiento arbustivo del maíz (“maize bushy stunt”), transmitido a plantas experimentales obtuvo un 60-80% de transmisión del fitoplasma. Otros cicadélidos, Macrosteles quadrilineatus y Scaphoides titanus, son eficientes vectores de los fitoplasmas del amarillamiento del aster y de la flavescencia, respectivamente. Existe también evidencia de que la infección del fitoplasma en el vector puede ser beneficiosa para ellos. Los insectos que portan el fitoplasma del enanismo arbustivo del maíz y del amarillamiento del aster presentan una mayor tasa de sobrevivencia que aquellos insectos libres de la infección de estos fitoplasmas, especialmente a bajas temperaturas, respaldando la hipótesis que los fitoplasmas pueden haber evolucionado en los insectos y secundariamente en las plantas hospedantes.
De manera interesante, los áfidos aparentemente no funcionan como vectores de espiroplasmas o fitoplasmas. Varias especies de áfidos se alimentan de las mismas plantas hospedantes y realizan una ingestión sostenida del floema, presumiblemente ingiriendo espiroplasmas y fitoplasmas como lo hacen su contraparte, los cicadélidos, pero nunca han mostrado que adquieran y/o transmitan molicutes. Es posible que los áfidos carezcan de la capacidad de llevar a su interior fitoplasmas en el conducto alimenticio o en las glándulas salivares o que los fitoplasmas sean incapaces de multiplicarse en la hemolinfa debido a respuestas inmunológicas en el insecto o la incapacidad de los molicutes de metabolizar carbohidratos en la hemolinfa del áfido. Sin embargo, ninguna de estas hipótesis ha sido experimentada empíricamente.
Epidemiología y Manejo
Las estrategias de manejo de las enfermedades causadas por espiroplasmas y fitoplasmas, pueden ser divididas en aquellas dirigidas al patógeno y aquellas dirigidas al vector. Los métodos para manejar al patógeno incluyen el uso de cultivares resistentes, los cuales sólo se conocen para unas cuantas especies de hospedantes. Un ejemplo de lo antes mencionado es el caso de la palma malaya enana, que se planta frecuentemente en áreas turísticas donde ha prevalecido anteriormente la enfermedad del amarillamiento letal del cocotero. Se han identificado también genotipos de fresno con tolerancia a los amarillamientos. Los bactericidas que inhiben la formación de la pared celular, como lo es la penicilina, presentan una acción nula contra estos molicutes sin pared celular, pero las tetraciclinas y otros productos que influyen en la fisiología de dichos procariotas pueden limitar la concentración del patógeno en la planta (McCoy 1982). Los productos relacionados con la tetraciclina han sido utilizados para preservar especímenes de árboles y arbustos en arreglos paisajísticos o jardines, pero no son una opción viable para cultivos debido a su costo y los residuos de éstos en productos alimenticios. Los molicutes han sido eliminados de ciertos materiales vegetales valiosos mediante el uso de terapias de calor o de CO2, o mediante el cultivo de meristemas. Las opciones de manejo dirigidas a los insectos vectores son difíciles de implementar debido a que un control efectivo sólo puede ser alcanzado sin las aplicaciones preceden a la migración de los insectos al campo. Si los productos químicos son aplicados después de la detección de los insectos en el cultivo, éstos tienen la capacidad de introducir el patógeno dentro de la planta antes que el productor se percate de la presencia del insecto. Los programas efectivos para amarillamiento del áster en zanahorias y lechuga se basan en el monitoreo de las migraciones de M. quadrilineatus, el principal vector de este fitoplasma en Estados Unidos.
Bacterias Fastidiosas Colonizadoras del Floema
Historia
Un subgrupo de enfermedades en plantas caracterizadas por síntomas tipo “amarillamientos” (clorosis foliar, enanismo, falta de vigor y muerte) y además necrosis del floema, visible como un anillo oscuro en secciones de tallos recién cortadas, son causadas por bacterias con pared celular. Estas enfermedades han presentado una serie de retos en su diagnóstico y manejo. La incapacidad, de manera consistente, de cultivar a estos microbios patógenos a partir de plantas afectadas ha llevado erróneamente a indicar a estos agentes causales como virus primero y posteriormente, una vez que los molicutes fueron caracterizados, como fitoplasmas. Aunque la mayoría de estos agentes causales aún presentan dificultades para su cultivo en medios artificiales, nuestro conocimiento de su naturaleza y diversidad ha aumentado considerablemente debido a los métodos recientes de caracterización molecular.
Biología
Las bacterias que habitan en el floema son generalmente bacilos muy pequeños que presentan una morfología celular procariota Gram negativa. La membrana que las cubre algunas veces es ondulada, una característica que le dio su nombre inicial de Rickettsia ("Rickettsia-like organism", en inglés o RLO). A medida que los científicos alrededor del mundo tienen mayor acceso a las herramientas moleculares y físicas existentes, tales como la microscopia electrónica, análisis de la secuencia 16S rADN y PCR, crece el número de enfermedades atribuidas a estas bacterias con pared celular colonizadoras del floema. En una revisión reciente, Davis (2001) listó veinticuatro enfermedades en monocotiledóneas y dicotiledóneas, plantas herbáceas y árboles, vegetales, frutales, granos y ornamentales. Estas enfermedades han sido citadas en los Estados Unidos, Canadá, la región del Caribe, Centro y Suramérica, Europa, Asia, África, el Medio Oriente y los países del Mediterráneo.
Solamente algunas bacterias fitopatógenas habitantes del floema han sido ubicadas taxonómicamente y se encuentran dentro del Phylum Proteobacteria. Candidatus Liberibacter asiaticus y Candidatus L. africanus, agentes causales del “Enverdecimiento de los Cítricos” (“Citrus greening” o Huanglongbing) en Asia y África, respectivamente, se asemejan al subgrupo Proteobacteria α-2. El agente no cultivable del crecimiento arbustivo de la papaya (“Papaya Bunchy Top”) se ubica dentro del subgrupo K-1 de la misma familia. El patógeno de una enfermedad de las cucurbitáceas recientemente reconocida y económicamente importante, denominada zarcillo amarillo (“Yellow Vine”) (figura 13), fue identificado recientemente como Serratia marcescens (Sm), un miembro de y-3 Proteobacteria. A diferencia de las otras bacterias colonizadoras del floema, Sm es fácilmente cultivada en medios bacteriológicos comunes y las diferentes cepas que infectan a las cucurbitáceas han mostrado diferencias significativas con las cepas aisladas de otros nichos ecológicos. S. marcescens es un microbio cosmopolita en productos alimenticios, desechos almacenados, cuerpos de agua y suelo; a menudo es un saprófito inocuo, pero puede causar enfermedades en vertebrados inmunocomprometidos, incluyendo a los humanos, y especies de insectos. Las cepas de Sm habitan en la rizosfera (zona radicular) de las plantas, produciendo efectos benéficos como rizobacterias promotoras del crecimiento (“Plant Growth Promoting Rhizobacteria”, - PGPR). Debido a ciertas propiedades antifúngicas, algunas cepas de Sm están siendo evaluadas como potenciales agentes biocontroladores en la agricultura. Sin embargo, ciertas cepas invaden los tejidos de las plantas y causan enfermedades económicamente devastadoras en cultivos de importancia agronómica.
 |
| Figura 13. |
Dado que la mayoría de las bacterias con pared habitantes del floema no son cultivables en medios artificiales, la mayoría de nuestro conocimiento de este grupo de patógenos respecto a ubicación filogenética proviene de los análisis moleculares de regiones clonadas tal como el 16S rADN. Patógenos de otras enfermedades importantes, tal como la hoja mazuda del trébol (“Clover Club Leaf”) no han sido caracterizados taxonómicamente. Sin embargo, se han utilizado otras herramientas para caracterizar a las bacterias con pared habitantes del floema. La bacteria agente causal del crecimiento meristemático arbustivo de la papaya (“Papaya Bunchy Top”) un bacilo de 0.25-0.35 por 0.8-1.6 µm, está rodeada de un tipo de pared Gram negativa que aparentemente carece de una capa de peptidoglicano en el espacio periplásmico. Las dos especies de Candidatus Liberibacter, aunque similar una a otra, causan diferentes síntomas en las especies de cítricos afectadas y se ven influenciadas de diferente manera por la temperatura ambiental.
Interacción Hospedero - Patógeno
Hasta la década de 1980, la enfermedad bacteriana degenerante del floema más estudiada era la hoja mazuda del trébol. Esta enfermedad se denominó de esa manera debido a que las hojas nuevas de Trifolium incarnatum (trébol rojo), se retardaban en abrir y permanecían enrolladas en forma de mazo. Este patógeno infecta a un amplio rango de hospedantes experimentales en por lo menos siete familias botánicas y otros síntomas incluyen a una ligera virescencia de las flores. Extractos obtenidos de plantas infectadas contienen cadenas de células (“talos”) que poseen motilidad ondulatoria que es distintiva de este grupo. Una enfermedad económicamente más importante es el “Enverdecimiento de los Cítricos”, que causa pérdidas severas en Asia y África (da Graca 1991). Los síntomas más prominentes de esta enfermedad incluyen clorosis de las nervaduras y ocasional moteado de las hojas, además de reducción del tamaño de las hojas y pecíolos erectos. Los frutos son asimétricos, miniaturas, de sabor amargo y se caen prematuramente. El nombre de la enfermedad “enverdecimiento” deriva del color de los frutos que permanecen en el árbol, que no maduran en el período normal de tiempo (Figura 14). El amarillamiento de zarcillos en cucurbitáceas, una enfermedad relativamente nueva, descrita al final de la década de 1980, causa pérdidas devastadoras en la producción de sandía, calabacín, zapallo y melón tipo “cantaloupe”, pero nunca ha sido citada afectando al pepino en el campo. Las plantas afectadas por esta enfermedad se tornan amarillas y marchitan rápidamente causando un colapso total de la planta. La necrosis del floema es evidente con una decoloración de tonalidad chocolate miel. Originalmente se pensó que esta enfermedad estaba confinada a la región del sur de Oklahoma y norte de Texas, pero recientemente se la ha identificado en varios estados del medio oeste y hacia el este en Tennessee.
 |
| Figura 14. |
Diseminación
Como es el caso para los molicutes, la taxonomía de los insectos que transmiten a las bacterias limitadas al floema es diversa. Los agentes causales del enverdecimiento de los cítricos (L. asiatum y L. africum) son transmitidos por psílidos, Trioza erytreae (Enverdecimiento Asiático) y Diaphorina citri (Enverdecimiento Africano), respectivamente. La bacteria puede ser adquirida en 30 minutos, pero su transmisión requiere de un período de latencia de 7-10 días (De Graca 1991), indicando ésto que se multiplica dentro del psílido vector.
La relación existente entre las bacterias habitantes del floema con respecto a las chicharritas vectoras ha sido menos estudiada. Los vectores del crecimiento meristemático arbustivo de la papaya incluyen al cicadélido Empoasca stevensii, aunque la naturaleza de la relación entre el patógeno y su vector es desconocida. El organismo causante de la hoja mazuda del trébol es transmitido por el cicadélido Agalliopsis novella, más no así por otros relacionados muy cercanamente al primero como son Agallia constricta y Aceratagallia sanguinolenta. Esta bacteria se multiplica dentro del vector en una relación propagativa.
Recientemente el agente causal del amarillamiento de los zarcillos de las cucurbitáceas, S. marcescens, se encontró que era transmitido por la chinche del calabacín, Anasa tristis (Figura 15). Esta relación es poco usual debido a que las chinches por lo general no se identifican como vectores de patógenos asociados con el floema de las plantas. La chinche del calabacín puede adquirir la bacteria luego de 24 horas, teniendo la capacidad de transmitirla de 1 a 21 días después de adquirirla. La bacteria está presente en la hemolinfa de algunos, pero no de todos los transmisores, lo cual es consistente con una relación no circulativa (se ubica en la parte anterior del intestino, no el hemocele) entre la bacteria y su vector.
 |
| Figura 15. |
Control
El uso de antibióticos para reducir o eliminar a los patógenos residentes en el floema ha tenido un éxito limitado. En una investigación respecto al manejo de la enfermedad del enverdecimiento de los cítricos, las inyecciones o las aspersiones de tetraciclina redujeron los síntomas foliares y en los frutos en el orden de 10-97% (da Graca 1991), pero otros antibióticos como la penicilina, fueron menos efectivos. La termoterapia (tratamiento con calor) puede ser efectiva, con hasta un 100% de control, cuando se someten las yemas de árboles infectados a una temperatura de 47 ºC por cuatro horas. Sin embargo, este método no es considerado práctico para su uso a gran escala (da Graca 1991). No existen tratamientos prácticos disponibles para plantas herbáceas afectadas por estas bacterias.
La reducción de la diseminación de patógenos mediante el uso de variedades resistentes al vector brinda la mejor oportunidad para un control a largo plazo de estas enfermedades que afectan el floema. Sin embargo, en el caso de infestaciones agudas del vector, el uso de insecticidas es la estrategia de manejo más efectiva. El uso de enemigos naturales y el impacto del uso de insecticidas en los parasitoides y depredadores, también debe ser tomado en consideración para las estrategias de manejo a largo plazo.
En estudios del amarillamiento de los zarcillos en cucurbitáceas, la exclusión del vector (Anasa tristis) de las plantas susceptibles y el uso de cultivos trampa para prevenir o reducir la infestación de este insecto dio como resultado una reducción de la incidencia de la enfermedad. Sin embargo, el mecanismo de control más efectivo de esta enfermedad es la aplicación de insecticidas para reducir las poblaciones del vector, la chinche del calabacín, Anasa tristis. Existe algún tipo de progreso en cuanto al desarrollo de variedades de sandía resistentes a la enfermedad causada por S. marcescens.
Historia
Las bacterias patógenas con pared celular también se localizan en las células transportadoras de agua y nutrientes del xilema de sus hospedantes y son transmitidas por chicharritas (“sharpshooters” Hemiptera, Cicadellidae) y por salivazos (“spittlebugs” Hemiptera, Cercopidae) que se alimentan del xilema (Raju and Wells 1986). Como en otros casos, se asumió en un principio que estas enfermedades eran causadas por virus, posteriormente se las denominó como “fastidiosas” debido a la dificultad de ser cultivadas en medios sintéticos y “parecidas a rickettsias” ("rickettsia-like"), debido a la similitud superficial de sus paredes rugosas con las de este grupo de organismos. Las Bacterias Fastidiosas Limitadas al Xilema (XLB, por sus siglas en inglés) incluyen ahora a un grupo de miembros que son cultivados en medios sintéticos especiales que causan varias enfermedades en un amplio rango de hospederos. Clavibacter xyli subsp. xyli (Raquitismo de la Soca en Caña de Azúcar) y Clavibacter xyli subsp. cynodontis (Atrofiamiento del Pasto Bermuda), dos ejemplos de bacterias limitadas al xilema, son bacterias bacilares fastidiosas de crecimiento lento en medio de cultivo, Gram positivas, de variadas formas que a menudo pueden presentarse como corineiformes (en forma de mazo) y a menudo se presentan en pares unidos en un extremo, adquiriendo la forman de una “V” (Davis and Vidaver 2001). Otras formas Gram-positivas incluyen a varios patovares de Curtobacterium flaccumfaciens que causan enfermedades vasculares en remolachas, frijoles,soya, tulipanes y flor de Pascua (Poinsettias) (Davis and Vidaver 2001).
Xylella fastidiosa, la más estudiada de las Bacterias Fastidiosas Limitadas al Xilema (XLB), presenta un amplio rango de hospederos y causa una afección severa en la mayoría de sus hospedantes. La enfermedad de Pierce de la vid ha incidido en California por muchos años, y recientemente ha sido identificada en Europa. La quemazón foliar de la adelfa o laurel rosa (Nerium oleander), almendros y varios árboles forestales, “Phony peach” del duraznero, y el escaldado de la hoja en ciruelas son ejemplos de enfermedades causadas por esta bacteria en California, Georgia y otros estados de Estados Unidos. En Taiwán, la quemazón de la hoja del peral se le atribuye a X. fastidiosa, y en Brasil y Argentina, X. fastidiosa causa la clorosis variegada de los cítricos (Figura 16) la cual se ha tornado en una limitante de la producción de cítricos y el mismo patógeno fue identificado como el agente causal de un escaldado de la hoja en cafeto. En cada una de estas enfermedades la bacteria es transmitida de planta a planta por insectos vectores, particularmente miembros del suborden Auchenorrhyncha, de la familia Cicadellidae (“sharpshooters”). La enfermedad de Pierce de la vid se consideró al inicio como una enfermedad de menor importancia en California, debido a que su vector era relativamente sedentario. Sin embargo, esta enfermedad ha tomado un papel más devastador recientemente, amenazando a las industrias de las uvas y del vino en dicho estado. El cambio en la importancia de esta enfermedad proviene de la invasión del área dedicada al cultivo de uvas de Homolodisca coagulata (figura 17). un miembro de la familia Cicadellidae. Dicha importancia se dio solamente por este factor sin que ocurriera una mutación en los genes de virulencia de este patógeno bacteriano ni de la ampliación en la siembra de cultivares de uva susceptibles. El insecto adquiere a X. fastidiosa a medida que se alimenta en los viñedos del sur y centro de California, movilizándose rápidamente de allí a grandes distancias, alimentándose a medida que pasa por ellas y diseminando eficientemente a la bacteria de una planta a otra y de viñedo en viñedo.
 |
 |
| Figura 16. |
Figura 17. |
Biologia
X. fastidiosa es un bacilo pequeño, angosto, Gram-negativo que crece lentamente en medios especiales (Hopkins 2001). Las cepas y patotipos de la especie han sido identificados y caracterizados mediante el uso de varios métodos moleculares que incluyen: rep-PCR, RAPD-PCR, RFLP e hibridación ADN-ADN. La relación hospedero patógeno que conlleva a la expresión de los síntomas de la enfermedad no esta completamente dilucidada, pero lo más probable es que una población elevada de la bacteria ocluya los vasos xilemáticos, lo que conduce a la muerte de la planta afectada. Xylella fastidiosa fue el primer fitopatógeno bacteriano cuyo genoma se secuenció completamente, este trabajo fue llevado acabo por un equipo de científicos brasileños que trabajaron en conjunto como un consorcio. Otros científicos han derivado una información valiosa de dichos estudios, tanto de la propia secuencia del genoma como de su alineamiento con el de otros microorganismos (Dow and Daniels 2000). Sin embargo, estos notables logros no han dado como resultado el desarrollo de un sistema genético por medio del cual se estudie las características de patogenicidad, pero los plásmidos nativos aislados y caracterizados de varias cepas de la bacteria pueden ser útiles como vectores para su transformación bacterial.
Interacción Hospedero-Patógeno
Acerca de X. fastidiosa se pensó originalmente que sólo infectaba a unas cuantas plantas leñosas, que incluían la vid. A medida que la tecnología avanzó, más enfermedades de etiología desconocida fueron atribuidas a X. fastidiosa, y el rango de hospederos incluye ahora a más de 75 plantas, algunas de las cuales se listan en el (cuadro 1).
Cuadro 1. Algunas plantas hospederas y enfermedades causadas por X. fastidiosa.
| PLANTA |
ENFERMEDAD |
| Vid |
Enfermedad de Pierce |
| Cítricos |
Clorosis variegada de los Citricos |
| Adelfa, laurel rosa |
Quemazón foliar |
| Almendro |
Quemazón foliar |
| Roble |
Quemazón foliar |
| Sicamoro |
Quemazón foliar |
| Alfalfa |
Enanismo de la Alfalfa |
| Duraznero, Melocotonero |
“Phony peach” |
Los síntomas de la enfermedad de Pierce de la vid puede ser detectada de 3 a 15 meses después de la infección inicial y usualmente incluye 1) escaldado, secamiento progresivo de las hojas de los márgenes hacia el pecíolo de la hoja, 2) se tornan chocolates las áreas escaldadas, 3) enanismo, marchitamiento o desecamiento de parte o toda la parra o vid, 4) muerte gradual del sistema radicular, 5) muerte de la vid en un periodo de tres a cuarenta y ocho (3-48) meses (figura 18). En árboles de naranja dulce, los síntomas de clorosis variegada incluyen a zonas conspicuas de variegación en el haz de las hojas viejas y lesiones levantadas en el envés de la hoja. Los frutos son pequeños y duros. La quemazón foliar de árboles en jardines y de frutales típicamente presenta una necrosis internerval y marginal irregular que se expande a lo largo del limbo foliar, causando deformaciones y defoliaciones prematuras. Muchos hospederos pueden ser asintomáticos.
 |
| Figura 18. |
Los síntomas asociados con la infección de X. fastidiosa sugieren que la conducción de agua y nutrientes del xilema es interrumpida por la presencia de la bacteria. Los agregados bacterianos, los depósitos de goma o las tilosis que bloquean los elementos conductores, todos ellos contribuyen a los estreses de agua que presentan las plantas infectadas. Como dato interesante, microfotografías electrónicas de plantas afectadas han mostrado variaciones en cuanto al número y tipo de oclusión de los elementos del xilema, lo que hace que se presenten discrepancias en cuanto a los mecanismos del desarrollo de la enfermedad. Una teoría indica que una fitotoxina liberada por el patógeno es responsable por la quemazón foliar. Otra posibilidad es que la infección causada por X. fastidiosa afecte el balance de los reguladores de crecimiento. Se ha aplicado ácido giberélico a árboles de melocotón afectados por “Phony peach” mostrando algún tipo de remoción de los síntomas.
Diseminación
X. fastidiosa es transmitida por homópteros que se alimentan del xilema. Este grupo de insectos incluye a chicharritas “sharpshooters” (Hemiptero, subfamilia Cicadellinae), y a los saliveros (Hemiptero, Familia Cercopidae). Es posible que las cicadas (Hemiptero, familia Cicadidae), que también se alimentan del xilema, puedan transmitir a X. fastidiosa, pero ésto todavía no ha sido demostrado.
Varias especies de chicharritas del tipo “sharpshooters” son vectores naturales o experimentales de X. fastidiosa en cuanto a su interacción con la vid, pero sólo tres son considerados como de importancia económica: Draeculacephala minerva, Graphocephala atropuntata y Homalodisca coagulata. De estas tres especies, Homalodisca coagulata es considerado el más peligroso de los vectores debido a su introducción reciente y proliferación rápida en las áreas de los viñedos y cítricos en California.
Aunque Homalodisca coagulata previamente fue un vector menor de X. fastidiosa, el rango de hospederos de este insecto recientemente ha mostrado que incluye no sólo a la vid, Nerium oleander y cítricos sino a un numeroso grupo de plantas leñosas. Este insecto es capaz de sobrevivir, bajo ciertas condiciones, el periodo de invierno y portar a X. fastidiosa por periodos prolongados de tiempo. Además es capaz de alimentarse de tejidos leñosos en estado de dormancia, de los cuales también puede adquirir al patógeno.
La relación existente entre X. fastidiosa y su vector es inusual. La bacteria es adquirida por el insecto al alimentarse de tejido infectado y coloniza el canal alimenticio dentro del estilete, el cual es utilizado por el insecto para punzar los tejidos de la planta. La bacteria adherida a la cutícula del insecto crece formando una película fina que se alinea en la superficie interna del canal alimenticio. Los adultos y ninfas son capaces de transmitir a X. fastidiosa inmediatamente después de su adquisición, pero las ninfas pierden la capacidad de transmisión al mudar. Un adulto que adquiere a X. fastidiosa puede retener su capacidad de transmitirla por varios meses a toda la vida del insecto.
Epidemiología y Manejo
X. fastidiosa se considera como uno de los fitopatógenos que esta emergiendo a nivel mundial debido a su potencial impacto económico en dos de las más importantes materias primas agrícolas como lo son las uvas y los cítricos. En un menor grado también afecta a la industria de la producción de ornamentales. El manejo de este patógeno se enfoca en dos frentes: control del vector y desarrollo de variedades resistentes.
Las infestaciones agudas del vector de X. fastidiosa pueden ser controladas mediante tratamientos químicos de la planta hospedante; tanto los insecticidas de contacto como los sistémicos han demostrado su relativa efectividad en reducir las poblaciones de estos insectos. Sin embargo, desde hace tiempo la oposición pública al uso de pesticidas para erradicar insectos ha sido muy considerable. Las alternativas biológicas a los pesticidas sintéticos incluyen a avispas parasitoides y entomopatógenos, pero históricamente estos mecanismos no han sido efectivos para manejar a la enfermedad. El manejo de X. fastidiosa y sus vectores es un tema actual de investigación, especialmente en las regiones viníferas de California en donde la enfermedad de Pierce amenaza la salud económica de esta industria.
Las estrategias a largo plazo deben incluir el desarrollo de variedades resistentes, una perspectiva desafiante debido a que muchos de los hospedantes de X. fastidiosa son plantas perennes. Un enfoque novedoso a su control puede darse a través de la manipulación genética de las bacterias entéricas que viven naturalmente en el intestino de los cicadélidos. Mediante el uso del proceso de paratransgenesis, los investigadores esperan transformar estas bacterias que normalmente viven en el intestino de dichos insectos, para que secreten una toxina letal contra X. fastidiosa. La bacteria transformada pasaría de insecto a insecto mediante actividad coprófaga (ingestión de materia fecal) normal, afectando negativamente a X. fastidiosa a medida que es adquirida por los insectos paratransgénicos.
Conclusiones
Las bacterias habitantes del sistema vascular, con o sin pared celular, son un grupo diverso de fitopatógenos que a menudo mantienen una relación compleja e íntima con sus insectos vectores al igual que con sus plantas hospedantes. A pesar de su naturaleza “fastidiosa” o en algunos casos su patogenicidad obligada, las nuevas estrategias de investigación que no requieren de su cultivo han permitido progresos significativos en revelar los mecanismos de comportamiento y moleculares mediante los cuales estos procariotas invaden, colonizan e impactan el crecimiento y productividad de sus hospedantes. Estas áreas prometedoras de investigación podrán ciertamente facilitar el desarrollo de herramientas más efectivas de detección e identificación, al igual que resaltarán los puntos débiles en el ciclo de la enfermedad, lo que permitirá significativos avances en nuestra capacidad para moderar los efectos de estos organismos en nuestros cultivos alimenticios, de fibras y ornamentales.
Reconocimientos
Se desea agradecer a S. Von Broembsen y L. Littlefield por la revisión crítica del manuscrito. Esto es una publicación de Oklahoma Agricultural Experiment Station, Project # OKLO 2052.
Referencias
Calavan, E.C. and G.N. Oldfield. 1979. Symptomatology of spiroplasmal plant diseases. Pp. 37-64 in Whitcomb, R.F. and J.G. Tully, eds. The Mycoplasmas, Vol. III. Academic Press, Inc., New York.
da Graça, J.V. 1991. Citrus greening disease. Annu. Rev. Phytopathol. 29:109-136
Davis, M.J. 2001. Fastidious phloem-limited bacteria. Pp. 275-282 in Laboratory Manual for the Identification of Plant Pathogenic Bacteria; 3rd Edition. N.W. Schaad, J.B. Jones and W. Chun, eds., APS Press, Inc., St. Paul, MN
Davis, M.J. and A.K. Vidaver. 2001. Coryneform plant pathogens. pp. 218-235 In: N.W. Schaad, J.B. Jones and W. Chun. Laboratory Guide for Identification of Plant Pathogenic Bacteria. 3rd Ed. APS Press, Inc., St. Paul, MN.
Dow, J.M. and M.J. Daniels.2000. Xylella genomics and bacterial pathogenicity. Yeast 17:263-271.
Fletcher, J., A. Wayadande, U. Melcher and F. Ye. 1998. The phytopathogenic mollicute-vector interface: a closer look. Pytopathology 88: 1351-1358.
Hackett K.J. and T.B. Clark. 1989. Spiroplasma ecology. Pp. 113-200 in Whitcomb, R.F. and J.G. Tully, eds., The Mycoplasmas, Vol. 5. Academic Press, Inc.; New York.
Hopkins, D.L. 1989. Xylella fastidiosa: Xylem-limited bacterial pathogens of plants. Ann. Rev. Phytopathol. 27:271-290.
Hopkins, D. 2001. Xylella fastidiosa. pp. 201-213 In: N.W. Schaad, J.B. Jones and W. Chun, eds. Laboratory Guide for Identification of Plant Pathogenic Bacteria. 3rd Ed. APS Press, Inc., St. Paul, MN.
Lee, I.-M., R.E. Davis and J. Fletcher. 2000. The Mollicutes: Spiroplasmas and Phytoplasmas. In: N.W. Schaad, ed. Laboratory Guide for Identification of Plant Pathogenic Bacteria. 3rd Ed. APS Press, Inc., St. Paul, MN.
McCoy, R.E. 1982. Use of tetracycline antibiotics to control yellows diseases. Plant Dis. 66:539-542.
Raju, B.C. and J.M. Wells. 1986. Diseases caused by fastidious xylem-limited bacteria and strategies for management. Plant Dis. 70:182-186.
Razin, S., D. Yogev, and Y. Naot. 1998. Molecular biology and pathogenicity of mycoplasmas. Microbiol. and Molec. Biol. Rev. 62:1094-1156.
Sears, B.B. and B.C. Kirkpatrick. 1994. Unveiling the evolutionary relationships of plant-pathogenic mycoplasmalike organisms. Am. Soc. Microbiol. News 60:307-312.

