Schumann, G.L. and K.J. Leonard. 2000. Ferrugem do colmo do trigo (Ferrugem Negra do Trigo, pt). Portuguese translation by Sandra M. Mathioni e Eduardo Alves, 2013. The Plant Health Instructor. DOI: 10.1094/PHI-I-2013-0724-01. Updated 2011.
Ferrugem do colmo (ou ferrugem preta, ou negra, pt)
Puccinia graminis f. sp. tritici
Trigo e cevada, bérberis (uva-espim, pt) (e algumas outras espécies de Berberis, Mahoberberis e Mahonia spp.)
Authors
Gail L. Schumann, University of Massachusetts, Amherst
Kurt J. Leonard, U.S. Department of Agriculture,
Agricultural Research Service, Cereal Disease Lab, St. Paul, MN
Tradutores
Sandra M. Mathioni1 e Eduardo Alves2
1Plant Biology Division, The Samuel Roberts Noble Foundation, Ardmore, OK, USA
2Departamento de Fitopatologia, Universidade Federal de Lavras, Lavras, MG, Brasil

Uredinia de Puccinia graminis f. sp. tritici |
A ferrugem do colmo foi a doença mais temida dos cereais. Não é tão prejudicial atualmente devido ao desenvolvimento de cultivares resistentes, mas os surtos podem ocorrer quando novas raças do patógeno surgirem contra os tipos existentes de resistência as quais não são eficazes. A ferrugem do colmo continua a ser uma importante ameaça para o trigo e a cevada e assim, para o abastecimento mundial de alimentos. Anton deBary demonstrou pela primeira vez o ciclo de vida heteroécio do fungo da ferrugem, Puccinia graminis, o agente causal da ferrugem do colmo.
Sintomas
No trigo e em outras gramíneas hospedeiras:
As plantas geralmente não apresentam sintomas típicos da doença até 7 a 15 dias após a infecção, quando as pústulas ovais (uredínias), com urediniósporos de cor pardo-ferruginosa, rompem a epiderme (Figuras 1 e 2). Microscopicamente, esses esporos são equinulados e avermelhados (Figuras 3 e 4). As pústulas podem ser abundantes e produzidas em ambas as superfícies das folhas e dos colmos das gramíneas hospedeiras. Mais no final da estação de cultivo, as pústulas (télias) com teliósporos escuros começam a aparecer nas gramíneas infectadas (Figura 5). Microscopicamente, observa-se que os teliósporos são formados bicelulares, elipsoides e com paredes espessas (Figura 6).

Figura 1 |

Figura 2 |

Figura 3 |

Figura 4 |

Figura 5 |

Figura 6 |
No bérberis e em outros hospedeiros alternativos:
As pícnias aparecem em plantas de bérberis (Figura 7) na primavera, normalmente na superfície superior das folhas. Elas estão muitas vezes em pequenos grupos e exalam picniósporos em um exsudado pegajoso (Figura 8). Cinco a 10 dias mais tarde, estruturas em forma de taça cheias de aeciósporos laranja-amarelos e pulverulentos rompem a superfície inferior da folha (Figura 9). As aécias são amarelas e algumas vezes chegam a estender-se até 5 mm a partir da superfície da folha (Figura 10). Microscopicamente, observa-se que os aeciósporos têm uma superfície ligeiramente verrugosa (Figura 11).

Figura 7 |

Figura 8 |

Figura 9 |

Figura 10 |

Figura 11 |
Biologia do Patógeno
Os fungos causadores da ferrugem são parasitas obrigatórios. Na natureza, eles requerem tecido vivo do hospedeiro para o crescimento e reprodução; não podem sobreviver como saprófitas. Na ausência de tecido vivo do hospedeiro, eles sobrevivem como esporos. Na maioria dos fungos causadores da ferrugem, somente os teliósporos são adaptados para sobreviver fora do hospedeiro vivo por mais do que alguns meses em condições de campo.
O fungo Puccinia graminis é heteroécio. Esta palavra descreve fungos da ferrugem que requerem duas plantas hospedeiras não relacionadas, como o trigo e o bérberis, para completar o seu ciclo de vida. A Puccinia graminis é macrocíclica, produzindo todos os cinco tipos de esporos: basidiósporos, picniósporos (espermácios), aeciósporos, urediniósporos (uredósporos) e teliósporos. Anton deBary, em 1865, identificou pela primeira vez a natureza heteroécia do ciclo de vida, mas o papel de cada estágio dos esporos não foi completamente compreendido até que John Craigie, um cientista canadense (canadiano, pt), estudou o patógeno em 1927.
Embora a ferrugem do colmo seja causada por uma única espécie de fungo, Puccinia graminis, existe considerável variação genética dentro da espécie. Em 1884, Eriksson descobriu subespécies hospedeiro-específicas ou "formas específicas" do fungo. Cada forma específica é designada em latim como uma forma specialis ou "f. sp." Todas as formae speciales tem aparência idêntica, mas variam na gama de hospedeiros. O patógeno que causa a ferrugem do colmo do trigo (Triticum aestivum) é Puccinia graminis f. sp. tritici. Outras formae speciales incluem P. graminis f.sp. secalis, agente causal da ferrugem do colmo do centeio (Secale cereale) e P. graminis f.sp. avenae, agente causal da ferrugem do colmo da aveia (Avena sativa). Ambos, Puccinia graminis f. sp. tritici e P. graminis f.sp. secalis, causam ferrugem do colmo em cevada. Por volta de 1916, Stakman e outros determinaram que dentro de P. graminis f.sp. tritici existem subdivisões genéticas chamadas raças. Mais tarde, raças também foram encontradas dentro de outras formae speciales.
Ciclo de Vida e Epidemiologia da Doença
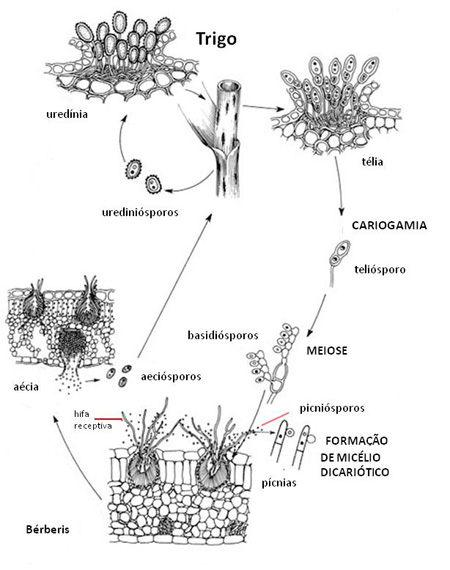
O ciclo da ferrugem do colmo do trigo começa com a exposição das culturas jovens de trigo aos esporos de Puccinia graminis f. sp. tritici, que são o inóculo primário. A fonte dos primeiros esporos que infectam as culturas jovens de trigo pode variar dependendo da região em que o trigo é cultivado. Em climas quentes, o trigo é plantado no final do outono e colhido no início do verão. Os primeiros esporos que infectam as plantas jovens de trigo são os urediniósporos. Estes geralmente vêm de plantas de trigo voluntárias e infectadas. Sementes espalhadas no campo ou nas estradas na época da colheita, muitas vezes germinam e produzem plantas voluntárias. Estas plantas podem ser infectadas a partir de esporos produzidos em plantas de trigo de maturação tardia ainda no campo. As plantas voluntárias e infectadas de trigo podem servir como uma ponte para o transporte de P. graminis f. sp. tritici durante o verão para a próxima safra de trigo que será semeada no outono.
Em regiões com clima temperado, o trigo pode ser plantado no outono (trigo de inverno) ou na primavera (trigo de primavera), dependendo da severidade do inverno (Figura 12). Por exemplo, algumas variedades de trigo de inverno podem sobreviver bem os invernos severos de Minnesota, Dakota do Norte e Manitoba, assim como a maior parte do trigo cultivado lá na primavera. Os primeiros esporos da ferrugem a infectar o trigo na primavera nas regiões temperadas podem ser aeciósporos de bérberis, o hospedeiro alternativo, ou urediniósporos das plantas de trigo infectadas em regiões distantes, com invernos mais amenos. Portanto, descrevemos dois ciclos da doença para a ferrugem do colmo – com ou sem bérberis.

Figura 12-map6a |

Figura 12-map6b |

Figura 12-map7a |

Figura 12-map7b |
Ciclo da doença com o bérberis
O bérberis é a fonte mais perigosa de inóculo primário da ferrugem do colmo em regiões temperadas. Se o bérberis cresce perto de campos de trigo, ele será uma fonte contínua de aeciósporos para as primeiras infecções do trigo na primavera Figura 13).
Figura 5 |

Figura 13 |
Puccinia graminis hiberna na forma de teliósporos escuros e com paredes espessas, diploides e que são produzidos no trigo ou em outras gramíneas hospedeiras mais para o final da estação de cultivo (Figura 5). A cariogamia (fusão de dois núcleos haploides para formar um núcleo diploide) e a meiose (divisão reducional para produzir quatro basidiósporos haploides) ocorrem nos teliósporos, os quais são produzidos na télia.
Na primavera, cada teliósporo germina para produzir basidiósporos haploides de paredes finas e incolores (Figura 14), os quais infectam os hospedeiros alternativos, como o bérberis comum.

Figura 14 |

Figura 15 |
Basidiósporos germinam e produzem micélio haploide que coloniza o tecido da folha. A partir deste micélio, são formadas pícnias no interior da folha, mas com projeções para o exterior que se estendem através da superfície e geralmente na superfície superior, nas folhas de bérberis. As pícnias produzem hifas receptivas e picniósporos (Figura 15). Nenhum desenvolvimento adicional ocorrerá até que as hifas receptivas da pícnia sejam fertilizadas por picniósporos de uma pícnia de outro grupo de acasalamento. Pícnias e picniósporos são também chamados de espermogônios e espermácias por alguns autores, mas os primeiros são os termos preferidos pelos especialistas em ferrugem.

Figura 16 |
Figura 8 |
Os picniósporos (Figura 16) são produzidos em uma substância pegajosa (melada) que é atrativa aos insetos, os quais ajudam a assegurar que a fertilização cruzada ocorra com sucesso (Figura 8). Os insetos por sua vez, carregam os picniósporos de uma pícnia para outra enquanto eles se alimentam da melada entre as folhas. Respingos de chuva também ajudam na dispersão dos picniósporos e na fertilização cruzada. A fertilização das pícnias é um ponto crítico no ciclo de vida do fungo da ferrugem pois dá origem ao micélio dicariótico. Depois de o núcleo do picniósporo se unir à hifa receptiva, os núcleos haploides emparelhados dividem-se em tandem no micélio ao longo das etapas restantes do ciclo de vida. Todas as infecções de ferrugem do trigo ou outras gramíneas envolvem esporos e micélios dicarióticos.
Ao longo dos dias, o micélio dicariótico cresce por toda a folha de bérberis até uma nova estrutura, a aécia, romper na superfície inferior da folha para liberar os aeciósporos dicarióticos (Figura 10). Os aeciósporos, embora produzidos em plantas de bérberis, infectam somente o trigo ou outras gramíneas que são hospedeiras de P. graminis. Os aeciósporos (Figura 11) diferem dos urediniósporos, que também infectam o trigo, na aparência - ligeiramente enrugados ao invés de espinhosos – e na forma em que são formados – em cadeias em uma aécia ao invés de hastes individuais em uma uredínia
Figura 10 |
Figura 11 |
No trigo, os aeciósporos germinam, os tubos germinativos penetram nas plantas e o fungo cresce como micélio dicariótico. Dentro de 1 a 2 semanas, o micélio de cada infecção produz uma uredínia com urediniósporos dicarióticos espinhosos e de cor vermelho-pardo, que rompem através da epiderme da folha ou do colmo (Figura 1).
Figura 1 |
Na ferrugem heteroécia, esta importante fase de esporos é chamada “fase repetitiva”, pois os urediniósporos são os únicos esporos da ferrugem que podem infectar a planta hospedeira em que foram produzidos. Em condições ambientais favoráveis, múltiplas e repetitivas infecções da mesma planta e das plantas vizinhas de trigo podem resultar em epidemias (epifitias, pt) explosivas.
Figura 5 |
Figura 6 |
Perto do final do ciclo da cultura, teliósporos escuros que hibernarão são formados nas télias (Figura 5), e o ciclo de vida está completo. Devido à cariogamia e à meiose ocorrerem nos teliósporos (Figura 6), este estágio dos esporos é muito importante pois é fonte de recombinação genética e também o seu papel como esporo de sobrevivência.

Figura 17 |
Ciclo da doença sem o bérberis (uva-espim, pt)
Na América do Norte, as epidemias (epifitias, pt) de ferrugem do colmo (negra, pt) podem ocorrer em regiões temperadas mesmo se o bérberis (uva-espim, pt) não estiver presente (Figura 17). Na ausência de bérberis (uva-espim, pt), os primeiros esporos de P. graminis a chegar no trigo da primavera são urediniósporos trazidos pelo vento e produzidos em culturas de trigo de inverno no Sul (Figura 18). O clima ameno ao longo da costa do Golfo do México permite que P. graminis sobreviva e se espalhe para áreas de trigo de inverno. Na primavera, os ventos predominantes no Sul, transportam os urediniósporos em direção norte para as Grandes Planícies (Great Plains) centrais onde infectam outras plantas de trigo de inverno. O clima nas planícies centrais é normalmente muito frio para permitir infecções da ferrugem do colmo durante o inverno. Quando o trigo de primavera começa a crescer no norte das Grandes Planícies, ele poderá ser infectado por urediniósporos trazidos pelo vento tanto do planalto central ou do sul das Grandes Planícies. No Norte, o ciclo da doença da ferrugem do colmo termina com a colheita do trigo.

Figura 18 |
No Sul, o ciclo da doença da ferrugem do colmo começa com urediniósporos que infectam plântulas de trigo de inverno após o plantio no outono. A maioria, se não todo, do inóculo primário é local. Ele vem de plantas voluntárias de trigo que brotaram e foram infectadas no verão. A disseminação de urediniósporos do Norte para o Sul é provavelmente menos importante. Trigo de primavera no Norte é colhido em agosto, muito antes da nova safra do trigo de inverno ter começado no Sul, onde o plantio não pode começar até outubro ou mais tarde. As plantas de bérberis (uva-espim, pt) não são infectadas no Sul, portanto não são um fator das epidemias (epifitias, pt) da doença naquele local. A razão disso é que os teliósporos de P. graminis não germinam a menos que forem expostos a longos períodos de temperaturas de congelamento.
Epidemiologia
A ferrugem do colmo é favorecida por dias quentes (25-30ºC/77-86ºF), noites amenas (15-20ºC/59-68ºF) e folhas molhadas de chuva ou orvalho. Aeciósporos e urediniósporos, ambos necessitam de água livre para a germinação, assim como os esporos dos outros estágios. As infecções ocorrem através dos estômatos (estomas, pt).
Figura 13 |
A fonte de inóculo pode ser predita a partir de um modelo da ferrugem. Se o inóculo provém de bérberis (uva-espim, pt), como fonte de inóculo, o padrão resultante da doença é geralmente em forma de leque, com o hospedeiro alternativo no ápice do leque (Figura 13). Se a doença tem um padrão mais uniforme, a fonte de inóculo é geralmente a partir de uma área ampla, como as culturas de trigo do sul (no hemisfério Norte) a partir das quais são liberados os urediniósporos. Infecções dispersas, principalmente sobre as folhas do topo de um campo de trigo, indicam que os esporos foram trazidos para o campo através do vento a partir de uma fonte externa. A chuva é importante para a deposição dos esporos durante a sua dispersão a longa distâncias.
Se a doença desenvolve-se em focos individuais dentro de um campo de trigo, a fonte de urediniósporos é provavelmente de micélio e/ou uredínia hibernados. Plantas doentes de focos originados de fontes de esporos hibernados, apresentam grande infecção em folhas mais baixas e menos infecção nas folhas mais jovens formadas no topo das plantas de trigo.
Na ausência de bérberis (uva-espim, pt) ou de outros hospedeiros alternativos, urediniósporos são os únicos esporos funcionais no ciclo da doença de P. graminis. Em climas tropicais e subtropicais, o micélio e os urediniósporos em plantas voluntárias de trigo e em outros hospedeiros é que começam as epidemias. Os urediniósporos geralmente são incapazes de sobreviver em condições de inverno rigoroso. No hemisfério Norte, o inóculo para o trigo de primavera chega das áreas do Sul. No hemisfério Sul, os urediniósporos chegam de áreas mais amenas do Norte. Ocasionalmente, P. graminis pode hibernar em plantas voluntárias de trigo, em outras gramíneas não hospedeiras e em trigo de inverno, mas geralmente apenas quando a neve cobre e isola ambos, as folhas de trigo e o micélio do fungo. Isso é mais provável de ocorrer quando o trigo de inverno é plantado diretamente no restolho de trigo da safra anterior.
Os urediniósporos são produzidos cerca de 7 a 15 dias após a infecção, por isso pode haver várias gerações de inóculo produzidos durante uma única estação de cultivo. Uma uredínia pode produzir pelo menos 100.000 urediniósporos. Epidemias (epifitias, pt) explosivas podem ocorrer durante condições ambientais favoráveis, resultando em perdas de 50% a 70% ao longo de uma região.
A ferrugem do colmo provoca perdas de rendimento de cereais de várias maneiras. O fungo absorve nutrientes dos tecidos da planta os quais seriam utilizados na formação de grãos em uma planta sadia. Como as pústulas rompem o tecido epidérmico, torna-se difícil o controle da transpiração pelas plantas, por isso o metabolismo se torna menos eficiente. Dessecação ou infecção por outros fungos e bactérias também podem ocorrer. A interferência que ocorre nos tecidos vasculares resulta em grãos murchos. A ferrugem do colmo pode também enfraquecer os colmos do trigo, resultando em acamamento (acama, pt) durante ventos fortes e chuva (Figura 19). Onde ocorrer severo acamamento (acama, pt), as culturas não poderão ser colhidas mecanicamente.

Figura 19 |
Manejo da doença (Meios de luta, pt)
Erradicação do bérberis (uva-espim, pt):
Uma vez que o ciclo de vida de P. graminis foi determinado, tornaram-se mais claros os efeitos potenciais da remoção do hospedeiro alternativo bérberis (uva-espim, pt) (Figuras 20 e 21). Um levantamento caro e extensivo sobre o bérberis (uva-espim, pt) (Figura 22) e um programa de erradicação foi iniciado em 1918 nos EUA (Figura 23) e continua de uma forma limitada até hoje (Figura 24).

Figura 20 |

Figura 21 |

Figura 22 |

Figura 23 |

Figura 24 |

Figura 25 |
Originalmente esperava-se que o programa iria eliminar a ferrugem do colmo de ser uma doença importante na América do Norte, porque os basidiósporos não teriam o bérberis (uva-espim, pt) para infectar, e urediniósporos não poderiam geralmente sobreviver em condições de inverno rigoroso. A importância da dispersão continental das epidemias (epifitias, pt) de ferrugem do colmo não foi entendida até mais tarde. Urediniósporos hibernam em campos de trigo nas regiões Sul dos EUA e norte do México e são, então, transportados por via aérea para o norte através do que é agora chamado o "Caminho da Puccinia" (Figura 25). Se o clima é favorável para o desenvolvimento da doença no Sul, os urediniósporos irão chegar em tempo e em número suficiente para causar epidemias nas regiões de cultivo de trigo no Norte.
Apesar deste problema, a erradicação de bérberis (uva-espim, pt) teve efeitos positivos e significativos no controle de epidemias (epifitias, pt) de ferrugem do colmo (negra, pt). Em primeiro lugar, a fonte de inóculo primária foi removida significativamente. Uma planta de bérberis (uva-espim, pt) pode produzir até 64 bilhões (milhares de milhões, pt) de aeciósporos. Em segundo lugar, a variação genética da população do fungo foi reduzida através da eliminação do ciclo sexual, deixando apenas os urediniósporos assexuados para manter o fungo. Mutação é agora a fonte principal de variabilidade genética. Consequentemente, não há mais tantas raças diferentes da ferrugem do colmo (negra, pt) do trigo contra as quais os melhoristas (melhoradores, pt) de trigo devem buscar resistência. E finalmente, as epidemias são atrasadas por várias semanas em muitas das grandes regiões produtoras de trigo nos EUA e Canadá, pois os aeciósporos foram liberados antes da primeira chegada de urediniósporos do Sul.
Práticas culturais (Luta cultural, pt)
Há muito que se sabe que a umidade nas folhas e o nitrogênio (azoto, pt) foliar excessivo favorecem as infecções por fungos causadores da ferrugem. Os agricultores consideram esses fatores através do espaçamento, orientação da linha e horários de aplicação de fertilizantes. As mudanças recentes nas práticas de produção podem ter efeitos sobre a ferrugem do colmo (negra, pt). Em algumas áreas, as culturas de trigo de verão são irrigadas, o que pode aumentar a sobrevivência de plantas voluntárias infectadas. Além disso, muitos agricultores estão utilizando o plantio direto ou plantio mínimo. Isso aumenta a probabilidade de que o fungo da ferrugem hiberne na camada protetora de palha da cultura anterior.
O uso de variedades de trigo com maturação precoce nas Grandes Planícies centrais dos EUA, tem ajudado a reduzir a ameaça de epidemias (epifitias pt) de ferrugem do colmo (negra, pt). Variedades modernas de trigo naquela região, maturam cerca de 2 semanas mais cedo do que as variedades mais antigas. Isso limita o período para o desenvolvimento de epidemias (epifitias pt) de ferrugem do colmo (negra, pt) nas Grandes Planícies centrais, assim como o número de urediniósporos que podem contribuir para epidemias (epifitias, pt) mais ao Norte.
Resistência genética (Luta genética, pt)
A resistência genética é o meio mais comumente usado e mais eficazes para controlar a ferrugem do colmo (negra, pt). O seu sucesso na América do Norte está diretamente relacionado com o reduzido número de raças presentes na população do fungo após o programa de erradicação do bérberis (uva-espim, pt). Como o financiamento para o programa foi reduzido nos últimos anos, os cientistas temiam que os arbustos restantes de bérberis (uva-espim, pt) continuariam a se espalhar para as áreas de trigo, para servir como fonte de inóculo e como um meio pelo qual o fungo poderia completar seu ciclo sexual. Além disso, os cientistas descobriram que, mesmo na ausência de bérberis (uva-espim, pt), não poderia se esperar que os genes de resistência utilizados atualmente permanecessem eficazes indefinidamente pois o aparecimento de novas raças do fungo continuam a surgir por mutação. Pelo menos 50 genes diferentes para resistência raça-específica (vertical) à ferrugem do colmo (negra, pt) foram identificados em trigo ou transferidos para o trigo a partir de cruzamentos com parentais silvestres de trigo. Nem todos estes genes de resistência são igualmente úteis. Muitos foram rapidamente descartados a partir de programas de melhoramento de trigo, pois raças virulentas que poderiam superar sua resistência foram encontradas como prevalentes na população do fungo. Outros genes pareciam ser amplamente eficazes quando usados pela primeira vez, mas novas raças virulentas do fungo apareceram dentro de alguns anos do uso generalizado da nova resistência.
Por razões que não são entendidas completamente, alguns genes para resistência raça-específica (vertical) da ferrugem do colmo (negra, pt) do trigo permaneceram altamente eficazes por muitos anos. O mais bem sucedido destes genes foi o Sr31, um gene localizado em um segmento de um cromossomo de centeio que foi transferido para o trigo através de um processo complicado de hibridização interespecífica. O trigo contendo o gene Sr31 rapidamente tornou-se popular no mundo inteiro, uma vez que, além do gene Sr31, o segmento do cromossomo de centeio também continha genes para alta produção de grãos, bem como outros genes para a resistência a outras ferrugens. Desde os anos 1980, as variedades de trigo com Sr31 foram amplamente cultivadas em quase todas as principais regiões produtoras de trigo em todo o mundo, exceto na Austrália. A eficácia do gene Sr31 foi tão grande que a ferrugem do colmo (negra, pt) do trigo foi reduzida para níveis quase insignificantes em quase todo o mundo em meados da década de 1990.
Recentemente, a resistência de Sr31 foi finalmente superada. Uma nova raça do fungo da ferrugem do colmo (negra, pt) do trigo, altamente virulenta para as variedades de trigo contendo o gene Sr31, foi encontrada em Uganda em 1999. A nova raça, provisoriamente designada Ug99, rapidamente dominou a população do fungo em Uganda e se espalhou para o Quênia e Etiópia onde causou grandes epidemias (epifitias, pt) . Dentro de poucos anos, Ug99 foi encontrada na África do Sul e no Iêmen, de onde se espalhou para o norte e para o leste até o Irã. Parece inevitável que Ug99 em breve invadirá o Punjab, na Índia, uma das principais regiões produtoras de trigo do mundo. Exemplos anteriores de dispersão a longa distância de fungos causadores da ferrugem incluem a disseminação de uma raça da ferrugem do colmo (negra, pt) do trigo, da África do Sul para a Austrália, a disseminação da ferrugem do café, da África para a América do Sul, e a disseminação da ferrugem-polisora do milho, da América Central para a África. Para piorar a situação, a linhagem Ug99 do fungo da ferrugem do colmo (negra, pt) ampliou sua virulência através de mutações que lhe permitem superar a resistência de pelo menos dois outros genes de resistência vertical que os melhoristas (melhoradores, pt) de trigo têm confiado para a proteção contra ferrugem do colmo (negra, pt) na América do Norte e em muitos outras partes do mundo.

Figura 26 |
Em resposta à ameaça iminente de epidemias (epifitias, pt) de ferrugem do colmo (negra, pt) do trigo em todo o mundo, um esforço internacional foi organizado em 2008 para reduzir a vulnerabilidade dos cultivos mundiais de trigo às ferrugens. A organização, o Borlaug Global Rust Initiative, é coordenada por uma equipe da Universidade de Cornell e inclui líderes de pesquisa de dois centros internacionais de pesquisa agrícola, CIMMYT e ICARDA, a Organização para a Alimentação e Agricultura das Nações Unidas (FAO), e do Serviço de Pesquisa Agrícola do Departamento de Agricultura dos EUA. Esforços primários estão concentrados no desenvolvimento e implantação de nova resistência efetiva e global à ferrugem do colmo (negra, pt) do trigo. A resistência vertical deve ser considerada a curto prazo, mesmo que sua durabilidade seja questionável. A combinação de dois ou mais genes eficazes de resistência vertical irá proporcionar maior chance de uma resistência mais duradoura. A longo prazo, no entanto, os melhoristas (melhoradores, pt) de trigo podem depender mais de genes aditivos e de menores efeitos de resistência parcial expressos principalmente em plantas adultas (por exemplo, resistência horizontal). Uma série de linhagens de trigo de alto rendimento com níveis moderados de resistência horizontal já foram desenvolvidas no Centro Internacional de Pesquisa de Trigo e Milho (CIMMYT). Estas linhagens avançadas estão sendo intercruzadas para produzir variedades de trigo melhoradas que combinam até quatro ou cinco genes de resistência horizontal para suprimir eficazmente as epidemias (epifitias, pt) de ferrugem do colmo (negra, pt). Para preservar estas combinações eficazes de genes de resistência horizontal, mesmo quando as epidemias de Ug99 diminuírem, será necessário identificar marcadores genéticos para cada um dos genes de modo que os melhoristas (melhoradores, pt) possam continuar a selecionar para a sua presença, mesmo na ausência de doença.
Controle químico (Luta química, pt)
Em algumas áreas aonde a pressão da doença é elevada, fungicidas são aplicados no trigo para controlar as doenças de ferrugem. Os fungicidas que inibem a síntese de esteróis [por exemplo, inibidores da biossíntese de esteróis (SBIs) ou inibidores da desmetilação (DMIs)] são particularmente eficazes, mas o custo de aplicação é geralmente proibitivo para uso rotineiro na maioria das áreas de cultivo de trigo nos EUA.
Abordagens potenciais para o manejo (a luta, pt)
Os urediniósporos infectam o trigo somente através dos estômatos (estomas, pt). Os cientistas estudaram como os urediniósporos germinados localizam os estômatos (estomas, pt) na superfície das folhas (Figura 27). Embora vários fatores estão envolvidos, o tubo germinativo é capaz de detectar as células guarda pelas suas dimensões físicas em relação ao das células epidérmicas. Uma vez que um estômato (estoma, pt) é encontrado, um apressório é produzido e a infecção então começa. No futuro, poderá ser possível criar um trigo resistente, que é resistente à infecção dos urediniósporos, pois ele terá os padrões das células epidérmicas que não serão reconhecidos pelo fungo.
|
| 
Figura 27 |
Importância Histórica
A ferrugem do colmo (negra, pt) é uma das principais doenças do trigo e da cevada e portanto, uma ameaça potencial para o fornecimento mundial de alimentos. O trigo é a cultura para alimentação mais cultivada no mundo, e a cevada é a sexta maior. Juntos, eles representam mais de 25% da oferta mundial de alimentos. Estima-se que mais de US $ 5 bilhões (milhares de milhão, pt) são perdidos anualmente devido às ferrugens dos cereais (ferrugem da folha (castanha, pt), ferrugem do colmo (negra, pt), e ferrugem estriada (amarela, pt)). As ferrugens dos cereais provavelmente têm sido problemáticas desde que as primeiras culturas de cereais foram cultivadas no Crescente Fértil. Esporos de P. graminis foram encontrados em sítios arqueológicos em Israel datando de 1300 a.C. Trigo, cevada e bérberis (uva-espim, pt) originaram-se no Crescente Fértil, e assim essa relação complexa do ciclo de vida da ferrugem do colmo (negra, pt) ter uma história antiga.
A ferrugem do colmo (negra, pt) do trigo era um problema sério na Grécia e Roma antigas. A doença foi observada e reconhecida já na época de Aristóteles (384-322 a.C.). Os antigos romanos sacrificavam animais vermelhos, como cães, raposas e vacas para o deus da ferrugem, Robigo ou Robigus, a cada primavera, durante o festival chamado Robigalia na esperança de que a safra de trigo seria poupada da devastação pela ferrugem (Figura 28). Este festival foi incorporado ao calendário cristão primitivo como o Dia de São Marcos ou Rogações no dia 25 de abril. Registros climáticos históricos sugerem que uma série de anos chuvosos, em que a ferrugem teria sido mais grave e as colheitas de trigo reduziram, podem ter contribuído para a queda do Império Romano.

Figura 28 |
Embora a natureza parasítica da ferrugem do colmo (negra, pt) não fosse conhecida até 1700, os agricultores na Europa haviam observado muito mais cedo que o bérberis (uva-espim, pt) estava de alguma forma ligado às epidemias de ferrugem do trigo. Leis que proibiam o plantio de bérberis (uva-espim, pt) perto de campos de trigo foram aprovadas pela primeira vez em Rouen na França em 1660.
Os cientistas italianos Fontana e Tozzetti independentemente forneceram as primeiras descrições detalhadas do fungo da ferrugem do colmo (negra, pt) do trigo em 1767. Persoon o nomeou Puccinia graminis em 1797. Em 1854, os irmãos Tulasne reconheceram que algumas ferrugens autoécias (um hospedeiro) poderiam produzir até cinco estágios de produção de esporos. Eles foram os primeiros a conectar os estágios pardo-ferruginoso (urediniósporos) e escuro (teliósporos) como sendo diferentes esporos do mesmo organismo, porém os demais estágios de P. graminis permaneceram um mistério.
Anton deBary ficou intrigado com a falta de infecção quando basidiósporos de P. graminis foram colocados em plantas de trigo. Utilizando-se da observação dos agricultores de que o bérberis (uva-espim. pt) aumentou a ferrugem do trigo, ele conseguiu inocular bérberis (uva-espim, pt) com basidiósporos e observou os outros estágios de esporos se desenvolverem no hospedeiro alternativo. Uma vez que a natureza heteroécia do ciclo de vida foi estabelecida, muitos outros fungos causadores da ferrugem foram descobertos como sendo heteroécios, e os seus hospedeiros puderam ser identificados.
As plantas de trigo e de bérberis (uva-espim, pt) foram trazidas para a América do Norte pelos colonizadores europeus. O bérberis (uva-espim) possui uma série de usos, incluindo um corante amarelo produzido da casca, compotas e vinhos dos frutos, cabos de ferramentas a partir da madeira e sebes espinhosas de rápido crescimento para ajudar a manter os animais. Como na Europa, os agricultores começaram a reconhecer a conexão entre bérberis (uva-espim, pt) e epidemias (epifitias pt) de ferrugem do colmo (negra, pt) no trigo. Leis sobre bérberis (uva-espim, pt) foram promulgadas em várias colônias da Nova Inglaterra em meados de 1700. No entanto, bérberis (uva-espim, pt) continuou a se espalhar com os agricultores pioneiros, que estavam se mudando para o oeste. Ele também se espalhou dos plantios das fazendas para cercas e pequenos bosques. Os arbustos de bérberis (uva-espim. pt) podem atingir 3 m (9 pés) de altura e produzir muitos frutos os quais são atraentes para os pássaros e animais que se alimentam deles e espalham as sementes.
Figura 22 |

Figura 29 |
Após a epidemia (epifitia, pt) devastadora de ferrugem do colmo (negra, pt) na América do Norte em 1916, um programa cooperativo estadual e federal de erradicação de bérberis (uva-espim. pt) foi criado em 1918 (Figura 22). Este programa foi parcialmente motivado pela preocupação com o abastecimento de alimentos durante a guerra. A "guerra contra o bérberis (uva-espim, pt)" foi formada e alistava a ajuda da população em geral através do rádio e de anúncios em jornais, panfletos de extensão, e estandes em feiras, instando-os a ajudar na destruição de bérberis (uva-espim, pt). Mesmo as crianças das escolas foram incentivadas a ajudar a encontrar locais onde existiam os arbustos de bérberis (uva-espim, pt) (Figura 29). De 1975 a 1980, o programa foi gradualmente voltando para as jurisdições de vários estados. A quarentena federal ainda é mantida contra a venda de bérberis (uva-espim, pt) suscetível à ferrugem do colmo (negra, pt) nos estados que faziam parte no programa de erradicação de bérberis (uva-espim, pt). Um programa para testes de bérberis (uva-espim, pt) foi estabelecido para garantir que apenas as espécies e variedades de Berberis,, como o ornamental e popular Berberis Japonês, que são imunes à ferrugem do colmo (negra, pt), serão cultivadas na área de quarentena.
Referências Selecionadas
USDA-ARS Cereal Disease Lab Website: Home page: http://www.ars.usda.gov/main/site_main.htm?modecode=36-40-05-00
Black Stem Rust Biology and Threat to Wheat Growers (from a presentation to the Central Plant Board Meeting February 5-8, 2001, Lexington, KY). Cereal Disease Laboratory website, University of Minnesota.
Introduction to cereal rusts: http://www.ars.usda.gov/Main/docs.htm?docid=9854
Barberry information: http://www.ars.usda.gov/Main/docs.htm?docid=9747
Bushnell, W.R. and A.P. Roelfs, 1984. The Cereal Rusts. Vol. 1. Origins, Specificity, Structure, and Physiology. Academic Press, Orlando.
Carefoot, G.L. and E.R. Sprott, 1967. Famine on the Wind. Rand McNally and Co., Chicago.
Cook, R.J. and R.J. Veseth, 1991. Wheat Health Management. American Phytopathological Society Press, St. Paul, MN.
Dubin, H.J. and S. Rajaram, 1996. Breeding disease-resistant wheats for tropical highlands and lowlands. Annual Review of Phytopathology 34:503-526.
Large, E.C. 1940. Advance of the Fungi. Dover Publications, New York.
Leonard, K.J. and L.J. Szabo. 2005. Stem rust of small grains and grasses caused by Puccinia graminis. Molecular Plant Pathology 6:99-111.
Littlefield, L.J. 1981. Biology of the Plant Rusts: An Introduction. Iowa State University Press, Ames.
McIntosh, R.A. and G.N. Brown, 1997. Anticipatory breeding for resistance to rust diseases in wheat. Annual Review of Phytopathology 35:311-326.
Peterson, P.D. (ed.) 2001. Stem Rust of Wheat: From Ancient Enemy to Modern Foe. APS Press. St. Paul, MN.
Roelfs, A.P. 1982. Effects of barberry eradication on stem rust in the United States. Plant Disease 66:177-181.
Roelfs, A.P. 1989. Epidemiology of the cereal rusts in North America. Canadian Journal of Plant Pathology 11:86-90.
Roelfs, A.P., and W.R. Bushnell, 1985. The Cereal Rusts. Vol. 2. Diseases, Distribution, Epidemiology, and Control. Academic Press, Orlando.
Roelfs, A.P., R.P. Singh, and E.E. Saari, 1992. Rust Diseases of Wheat: Concepts and Methods of Disease Management. CIMMYT, Mexico, D.F.
Ug99 References
Borlaug Global Rust Initiative: http://www.globalrust.org/
FAO website on the spread of race Ug99: http://www.fao.org/agriculture/crops/rust/stem/rust-report/stem-ug99racettksk/en/
Singh, R.P., D.P. Hodson, J. Huerta-Espino, Y. Jin, P. Njau, R. Wanyera, S.A. Herrera-Foessel, and R.W. Ward. 2008. Will stem rust destroy the world's wheat crop? Advances in Agronomy 98:272-309. (a pdf file of this reference can be found at http://ddr.nal.usda.gov/bitstream/10113/36520/1/IND44295123.pdf
Stone, M. 2010. Virulent new strains of rust fungus endanger world wheat. Microbe 5:423-428.http://www.microbemagazine.org/index.php/09-2010-home/2849-virulent-new-strains-of-rust-fungus-endanger-world-wheat

