Reviewed feature article
McBeath, J.H. 2002. Snow mold-plant-antagonist interactions: survival of the fittest under the snow. The Plant Health Instructor. DOI: 10.1094/PHI-I-2002-1010-01.
Jenifer Huang McBeath
Plant Pathology and Biotechnology Lab
Agricultural and Forestry Experiment Station
University of Alaska Fairbanks
Fairbanks, AK 99775-7200
ffjhm@uaf.edu
|

Fig. 1. Snow mold patches on winter wheat caused by Microdochium nivale and Myriosclerotinia borealis (click image for larger view). |
|
Snow molds are obligate or facultative psychrophilic (cold-loving) fungi that can attack many economic plants under a cover of snow (Fig. 1). The dark, humid environmental conditions favor the development of psychrophilic microorganisms which are able to grow at temperatures of 0°C and below and have a maximal growth rate at about 20°C (27) (Fig. 2). Snow molds infect snow-covered plants when the temperature at the snow-plant interface remains around 0°C. They are comprised of more than a dozen species and occur commonly in high latitude regions. Most snow mold fungi that have been investigated are pathogens of economically important plants such as conifers, winter cereals, turf grasses, and flowers. Major snow mold diseases of conifers are caused by Neopeckia coulteri on pine (2) Herpotrichia juniperi on juniper (2), and Phacidium infestans on pine (16). The most economically important snow mold fungi are Myriosclerotinia borealis (Bubak & Vleugel), Kohn (Fig. 3), Coprinus psychromorbidus Redhead & J.A.Traquire. Microdochium nivale Samuels and Hallet, Typhula incarnata Lasch ex Fr., Typhula ishikariensis Imai, and Pythium iwayami S. Ito. Hirane (10). These snow mold fungi inflict serious damage on winter cereals and turf grasses. In addition, Typhula ishikariensis also causes serious damage on tulip bulbs in Russia (8) and Canada (7). M. borealis was found to act as a saprophyte and/or a weak parasite on Japanese and Dutch irises and other perennial flowers (28). In Alaska, M. borealis, C. psychromorbidus, and M. nivale are prevalent. T. incarnata, T. ishikariensis, and Pythium species, economically important snow mold fungi of other temperate and sub-arctic regions, have not been found in Alaska.
|

Fig. 2. Snow mold study; sample collection during break-up (click image for larger view). |
|

Fig. 3. Mycelial mat of Myriosclerotinia borealis on winter wheat under the snow (click image for larger view).
|
|
Snow mold damage to winter cereals and grasses is generally governed by environmental conditions during the fall, depth and duration of winter snow cover, and availability of pathogen inoculum (3). In Alaska (latitude 54°N to 71.5°N), agricultural activities are located mostly in the sub-arctic region where the number of snow-bound days ranges from 170 to 210 per annum with a maximum soil temperature of 12 to 15°C, due to the short growing season and permafrost (grounds with permanently frozen subsoil) (Fig. 4). In Alaska and other regions of high latitude and elevations, snow molds are the most serious diseases of winter wheat and turf grasses (22, 39). Snow mold fungi infect host plants in late fall or early winter when the soil is not yet frozen. During the long winters (November through early March), snow mold fungi proliferate and spread in host tissues under the dark, humid conditions associated with a thick snow layer.
 |
|
Fig. 4. Ice lens under a soil layer seen in
July. Many Alaska soils are on permanently
frozen ground (permafrost) (click image
for larger view).
|
Impact of Snow Layer on Plants
In northern countries and high elevation regions, where winter is long and harsh, a deep, persistent layer of snow insulates the roots and crowns of winter cereals, grasses, forages, ornamentals and shrubs, protecting them from winter injuries caused by prolonged exposure to lethal and sub-lethal low temperatures (3). In Alaska, a difference of 15 to 25°C was observed between ambient and soil temperatures with a snow depth of 53 cm. For instance, when ambient temperatures were -15°C and -45°C, temperatures at the root zone were registered at 0°C and -20°C, respectively. The dark, humid and relatively mild environment provided by the protracted snow cover prevents photosynthesis and drastically reduces plant metabolism. It also predisposes plants to attack by snow mold fungi by increasing the contact of soilborne inoculum to plant tissues (34) and by depleting the carbohydrate reserves (15). Snow mold resistant cultivars tend to metabolize carbohydrates more slowly (41). A positive correlation was found between host tolerance to freezing injury and resistance to snow mold fungus, Myriosclerotinia borealis (1).
Interactions of Snow Mold Fungi and Host Plants

Fig. 5. Snow stained by chlorophyll leaching from macerated plant tissues, caused by Myriosclerotinia borealis (click image for larger view). |
|
Many snow mold fungi are capable of producing extracellular enzymes (25,26,35). Comparisons of pectinase and cellulase activities of six snow mold fungi -- M. borealis, M. nivale, a strain of C. psychromorbidus (sclerotial-forming low temperature basidiomycetes, or sLTB), T. incarnata, T. idahoensis, and T. ishikariensis -- were made for polygalacturanse (PG), pectin methylesterase (PME), pectin transeliminase (PTE), and carboxymethycellulase activities. Among the six snow mold fungi, M. borealis has the highest level of enzyme activities in PG, PME, and PTE. Enzymes of sLTB have the lowest PG activites but the highest carboxymethylcellulase activities. The lowest PME and PTE activities were found in enzymes of T. incarnata but the lowest carboxymethyl-cellulase activities were found in enzymes of T. ishikariensis (Mehdizadegan and McBeath, unpublished data). The extracellular enzymes are believed to facilitate the degradation of plant tissues. Few snow mold activities were detected when soil temperatures fell below -5°C. The peak snow mold activities on plants occur during spring break-up when the soil temperatures are around freezing and the soil-snow interface is saturated with melting snow. Resistance in winter cereals appears to be correlated with total content of carbohydrate as well as fructan (41). Snow mold fungi might have a decreased ability to metabolize fructan polymers compared with simple sugars. Winter cereals that maintain a higher proportion of soluble carbohydrate as fructan were less susceptible (10). Death of the plant is due to maceration of plant tissues (leaves, sheaths, and especially the crowns), leaching of plant cytoplasm, depletion of carbohydrate reserves, and desiccation (Fig. 5).
Important Snow Mold Diseases
Coprinus Snow Mold. Receding snow reveals the presence of circular or irregularly shaped snow mold patches (Fig. 6) with white to gray, woolly or oppressed mycelium of Coprinus psychromorbidus. Infected plants have rotted, water-soaked leaves or leaf lesions (Fig. 7). The lesions, or leaves, become pale brown when dry and have dark red-brown margins (31,33,29). Coprinus snow mold, also called cottony snow mold, is caused by sclerotial and nonsclerotial strains of Coprinus psychromorbidus. The nonsclerotial strains of low temperature basidiomycetes (LTB) are recognized only as snow mold based on white to gray colored mycelium that has clamp connections. The sclerotial strains are similar but they produce abundant small (1 to 3 mm wide) sclerotia that are white first and then mature to become brownish black, irregularly shaped, and flattened. The nonsclerotial strains tend to grow faster and with more of a cotton-like appearance than the sclerotial strains and produce patches that are somewhat larger and whiter than those formed by the sclerotial strains (31,33,29).
 |
|
 |
|
Fig. 6. Snow mold damage on golf greens caused by Coprinus psychromorbidus; Chena Bend Golf Club, Fairbanks, Alaska (click image for larger view). |
|
Fig. 7. Receding snow reveals the presence of snow mold patch on winter wheat caused by Coprinus psychromorbidus (click image for larger view). |
Protracted snow cover favors the development of this disease. Beneath the snow on frozen ground, the optimum temperature for development of LTB is -3°C. Despite the requirement for prolonged exposure to zero or sub-zero temperatures for plant parasitism, Coprinus snow mold fungi grow optimally in sterile growth media from 10 to 15°C (12). Some strands of C. psychromorbidus are capable of producing extra-cellular enzymes (25, Mehdizadegan and McBeath, unpublished data), which seems to be an integral part of the disease development.
In Canada, sclerotia of sLTB are formed primarily on leaf surfaces. In Alaska, where the temperature in winter is harsher, sclerotia are found at the inner layer of the sheath tissue. The discrepancy may be an indication of adaptation. Sclerotia serve as the survival structure of the sclerotial strains. The means of persistence for the nonsclerotial strains are not known (29,31,33).

Fig. 8. Snow mold patch on winter wheat observed at the time of break-up (click image for larger view). |
|
Snow Scald. Snow scald is caused by Myriosclerotinia borealis (syn. Sclerotinia borealis Bubak & Vleugel). Disease development is favored by prolonged periods of deep snow cover on frozen soil. Patches of grayish fungal mass appear as snow recedes (Fig 8). Leaves are initially water-soaked and covered with sparse gray mycelia and tan sclerotia. Later they become bleached to white and desiccated. The optimum temperature for M. borealis development is -2°C. Similar to Coprinus psychromorbidus, M. borealis can grow optimally in sterile growth media from 10 to 15°C (12).
Sclerotia of M. borealis are variable in size. They may be spherical, oval or flake-like and are 0.5 to 7.0 mm long (29,31,33) (Fig. 9). Under favorable conditions, apothecia of M. borealis are up to 5 mm in diameter and are borne on a stipe (stalk) up to 6 mm high (29,31). The snow scald pathogen relies primarily on ascospores produced from apothecia for primary inoculum. Under the arid environmental conditions in interior Alaska with rainfall less than 13 inches per annum, the occurrence of carpogenic germination (germinating through fruiting body) of M. borealis is very rare. Sclerotia myceliogenic germination (germinating vegetatively from the thick-walled resting body) is found to be the primary means of dissemination. Mycelia were found entering host plants mostly through stomata, but direct penetration through the cuticle has also been observed (23). M. borealis produces copious amounts of extracellular enzymes, especially the pectinolytic enzymes, which appeared to play a significant role in rapid decomposition of leaves, sheaths, and crown, and thus death of the plant (23,25,26) (Fig. 10).
|

Fig. 9. Comparison of sclerotia of Myriosclerotinia borealis treated (right) and untreated (left) with Trichoderma atroviride, a hyperparasite (click image for larger view). |
|

A |
|

B |
|

C
Fig. 10. Snow mold disease developed on winter wheat caused by Myriosclerotinia borealis under the snow: (A) dark brown colored lesions on the sheath at soil line (late October); (B) maceration of plant tissue observed on winter wheat (mid-March); (C) plant at extensive stage of decomposition (May) (click each image for a larger view). |
|
Typhula Blight. Typhula blight is also called gray or speckled snow mold because of the characteristic white to gray-white mycelial mat that is speckled with numerous sclerotia. In areas with protracted snow cover the most important diseases of perennial grasses and winter cereals grasses are caused by species of Typhula. Typhula blight occurs from cool temperate to boreal regions of the Northern Hemisphere, except in Alaska. In a survey of golf course superintendents, diseases caused by Typhula spp. were the second most damaging on turf grasses in Canada. Treatment of this disease may account for half of the chemical fungicide used on turf in this country (13).
Typhula blight is caused by T. incarnata and three biotypes of T. ishikariensis. The symptoms of Typhula blight are often first apparent at snow melt as circular areas of straw-colored patches from 5 cm to 1 m or more in diameter (31). The leaves in affected areas are matted together and may be covered by gray-white mycelia. As the grass dries, the mycelia disappear and the leaves become grayish to silvery white, brittle, and encrusted over the patch. Large areas of turf may be killed when conditions are optimal for disease development. More often, however, only leaves are killed and plant crowns produce new leaves during the spring (31). Small, hard, spherical sclerotia form on or in infected leaves. The sclerotia are 0.2 to 5.0 mm in diameter. They are white, pink, amber, or chestnut-colored when young and reddish brown to black when mature. The sclerotia are useful in separating Typhula blight from pink snow mold (13,31).
Typhula spp. survive as sclerotia during the summer. In late autumn, the sclerotia become exposed to wet, cool (optimum 10 to 18°C) conditions that favor germination. Sporocarp (clavula) formation occurs under diffuse light high in the ultraviolet spectrum. Under direct light or darkness, sclerotia germinate by producing mycelia instead. It is believed that the mycelia originating from sclerotia serves as the primary inoculum rather than basidiospores developed from the clavula. Typhula spp. grows very well at temperatures just above freezing (1 to 2°C). The disease is most severe when deep snow or heavy mulch covers a wet turfgrass canopy and unfrozen soil conditions persist for a long period. Application of high rates of nitrogen fertilizer before winter dormancy may favor the disease by promoting succulent leaf tissue.
Pink Snow Mold. Pink snow mold is so named because of salmon-pink colored mycelia and sporodochia which appear on grasses after the snow recedes (Figs. 1, 11, and 12). Microdochium nivale requires little or no snow cover to cause disease. In some cool, humid regions, damages caused by M. nivale can be observed year round and to nearly all grass species. M. nivale can attack all plant parts during wet, cool periods in autumn and spring. It is less virulent than other snow mold fungi at temperatures below 5°C. Under favorable conditions, patches of affected turf first appear as small water-soaked spots less than 5 cm in diameter; they then rapidly change color from orange-brown to dark brown and finally to light gray with brown or pink margins. The spots are usually confined to diameters of less than 20 cm. On low-cut turfs, ring-like patches may appear, and the outer perimeter of the patch may have a water-soaked, greenish black margin (Fig. 13).
|
 |
|
 |
|
|
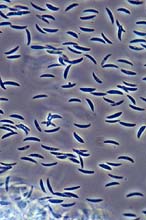
Fig. 11. Microdochium nivale spores (click image for larger view).
|
|
Fig. 12. Winter rye-otherwise same as Fig. 1 (click image for larger view).
|
|
|

Fig. 13. Snow mold patches on turf grasses caused by Microdochium nivale (click image for larger view). |
|
Microdochium nivale survives well in infected grasses and dead debris. When conditions are favorable, mycelia of the pathogen grow from infected debris or from germinating conidia to infected leaves. The disease is most severe when thatched turf is growing slowly in prolonged cool (0 to 8°C) and wet periods. Alternate thawing and snow cover, repeated frosts, cold fogs, and light drizzling rain result in particularly favorable conditions for leaf-to-leaf spread of the fungus. High nitrogen fertility, which promotes succulent grass, can increase turf susceptibility, whereas high levels of potassium tend to suppress the disease. Poor drainage and long leaf blades that become matted down, producing pockets of high humidity, also favor disease development. Spores and infected debris are easily carried on equipment wheels, mowers, animals, and shoes (29,31,33).
Snow Rot or Snow Blight. In cold weather, Pythium snow rot is well known on grasses in Japan and has been observed in Canada. This foliar blight is favored by high soil fertility, poor drainage, and saturated soil underlying deep snow. At least six Pythium spp. are known causal agents of snow blight. Small tan or orange spots may appear on the turf, or large areas of turf may be uniformly blighted. Rotted leaf tissue appears brown or light tan and is filled with oospores of the pathogen. Roots are mostly unaffected, but crowns can be rotted extensively. Plants with infected crowns die quickly (29,31,33).
Interactions among Snow Mold Fungi
The antagonism between snow mold fungi S. graminearum Elen and Typhula incarnata has been reported by Tomiyama (37). Competition for limited resources under snow has also created intraspecific competitive interactions among biotypes within T. incarnata, T. ishikariensis and LTB taxa (17,18). This highly competitive interaction among snow mold isolates has created a complex ecological structure whereby different survival and reproductive strategies exist among biotypes. Different vegetative incompatibility groups within biotypes of T. ishikariensis have adopted different survival strategies according to the prevailing cropping practices. Biological diversity among incompatible groups of T. ishikariensis decreases with increasing intensity of agriculture (20). Different vegetative incompatibility groups of snow molds tend to develop antagonistic relationships with each other on the periphery of individual patches as they compete for available resources (19).
In the interspecies interactions with complex infections of Pythium paddicum, Pythium iwayamai and T. incarnata, biomass of Pythium spp., especially P. paddicum, drastically decreased in the presence of T. incarnata, whereas T. incarnata biomass was seldomly affected by the presence of Pythium spp. T. incarnata appeared to be more competitive than Pythium spp. When T. incarnata coexisted with P. iwayamai, its population in leaves increased remarkably, suggesting the exploitation of Pythium spp., especially P. iwayamai by T. incarnata (36).
Interaction of Snow Mold and Antagonists
In Canada, Typhula phacorrhiza isolated by Schneider and Seaman was pathogenic to winter wheat. Applying a non-pathogenic strain of T. phacorrhiza at 500-2,000 kg/ha to creeping bentgrass, Burpee et al. successfully controlled Typhula blight comparable to the level achieved by chemical control using pentachloronitrobenzene (PCNB) applied at the rate of 30 kg/ha. Wu et al., studying 382 other isolates of T. phacorrhiza from corn soils in Ontario, Canada, confirmed the great variability in suppressive activity of these isolates but could not correlate it with their growth rates at 10°C. T. phacorrihza produces no antibiotics, nor is it parasitic on other Typhula spp. The mechanism of disease suppression has not been fully elucidated but laboratory studies indicate that competition for substrates and nutrients is a probable mechanism of suppression of T. ishikariensis and T. incarnata (4).
The low temperature fungus Acremonium boreale was also found to suppress T. ishikariensis, M. nivale, and C. psychromorbidus in Canada (32). Antibiosis is suspected to be the primary mechanism of disease suppression. A low temperature-tolerant bacterium, Pseudomonas fluorescens, was found to be antagonistic toward T. ishikariensis and was able to reduce snow mold damage (19).
Trichoderma atroviride, a cold tolerant, versatile hyperparasite isolated from the sub-arctic region of Alaska, was found to suppress a wide range of economically important plant pathogens, including C. psychromorbidus (Fig. 14) M. nivale, Myriosclerotinia borealis (Fig. 15), Pythium spp., T. incarnata (Fig. 15), T. idahoensis, and T. ishikariensis. T. atroviride is a mesophile, but well adapted to cold environments. It has a temperature range of 4°C (and less) to 33°C that makes it useful in controlling pathogens that can inflict damage on roots, stems, and other plant tissues under cool temperatures when plant tissues are particularly vulnerable. T. atroviride is fast-growing and produces profuse spores. It is rhizosphere competent and possesses plant growth promotion abilities (Fig. 18). T. atroviride CHS 861 is naturally resistant to metalyxyl (ridomil), captan, and pentachloronitrobenzene (PCNB or terraclor) (21,38).
|
 |
|
 |
|
|
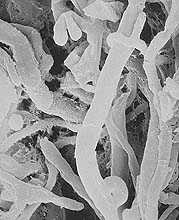
Fig. 14. Mycelium of Coprinus psychromorbidus showing typical clamp connection (parasitized by Trichoderma atroviride) (click image for larger view). |
|
Fig. 15. Hyphae of Typhula incarnata parasitized by Trichoderma atroviride (click image for larger view).
|
|
Trichoderma atroviride can utilize snow mold fungi as a food source. Hyphae of T. atroviride penetrated freely through the cell walls (Fig. 16) and became intertwined with the hyphae of the snow mold fungi (Figs. 15, and 16). Cell deterioration and lysis occurred rapidly. Chitinolytic enzymes produced by T. atroviride seemed to play a significant role in the mycoparasitism of T. atroviride against the snow mold fungi (5) (Fig. 16). In M. borealis, the sclerotia not only provide a means of survival but also serve as its primary source of inoculum. T. atroviride has been found to be capable of parasitizing the sclerotia of M. borealis and using it as a food source (21,38) (Figs. 9 and 17).
|
 |
|
 |
|
|
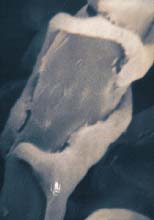
Fig. 16. Scanning electron microscopy of Coprinus psychromorbidus parasitized by Trichoderma atroviride in an advanced stage of deterioration (click image for larger view).
|
|
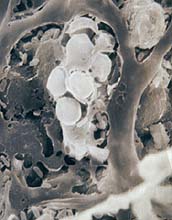
Fig. 17. Spores of Trichoderma atroviride formed in sclerotia of Myriosclerotinia borealis infected with Trichoderma atroviride (click image for larger view). |
|
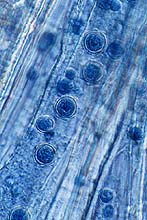
Fig. 18. Chlamydospores of Trichoderma atroviride in the root cells of winter wheat (click image for a larger view).
|
|
Efficacy studies of T. atroviride for the control of C. psychromorbidus were conducted under conditions of cold (about 4°C), dark and 100% humidity for 8 weeks. Because of the extreme conditions, a certain degree of mortality was observed in all treatments, including the blank control. No survivors (100% mortality) were found when winter wheat seedlings that were germinated from untreated seeds were challenged with C. psychromorbidus. A significant improvement in viability of winter wheat seedlings was observed when the seedlings were previously treated with T. atroviride biotype 603 (70% mortality) or CHS 861 (50% mortality). No difference was observed between uninfected plants treated with CHS 861 (39% mortality) and the blank control (39% mortality). Examination of the sheath of T. atroviride-treated winter wheat grown in snow mold contaminated soils revealed a dramatic reduction in the number of sclerotia formed by C. psychromorbidus. T. atroviride seemed to forge a symbiotic relationship with the roots of winter wheat plants (Fig. 18). T. atroviride mycelia, chlamydospores and chlamydospore primordia-like structures were observed in the root cells of winter wheat with no adverse effects on the plants (21,38).
Fairbanks Golf and Country Club is the golf club farthest north in the U.S. In a trial conducted there in September 2000 through May 2001, T. atroviride controlled snow mold fungi. Snow mold damage sustained by turf at the green ranged from 28 to 68% with a mean of 44.8%. No visible snow mold damage was found on turf grasses grown in adjacent areas where appropriate dosages of T. atroviride were applied (21) (Figs. 19 and 20).
 |
|
 |
|
Fig. 19. Trichoderma atroviride treated golf green (foreground) as seen in early May. Little or no snow mold damage was found on golf green. The untreated bunker (background) sustained heavy damage from snow mold infestation (click image for larger view).
|
|
Fig. 20. Golf green treated with recommended dosage of Trichoderma atroviride (left) compared to untreated area (right) (click image for larger view).
|
Conclusions
Some snow molds remain dormant through the summer and spend the winter in a vegetative state. Some other psychrotrophic molds can grow in summer without passing through a dormant stage (14). Beneath the snow on frozen ground, the optimum temperature for development of LTB is -3°C; for M. borealis it is -2°C. Despite the requirement for prolonged exposure to zero or sub-zero temperatures for plant parasitism, most snow mold fungi grow optimally in sterile growth media from 10 to 15°C (12). Conversely, M. nivale has an optimum in vitro growth temperature of 25°C, and can develop on winter cereals at all stages of plant growth (10). This discrepancy between the optimum growth temperatures of snow mold fungi under sterile and non-sterile conditions has been attributed to a poor competitive ability with other soil microorganisms (18). The adaptation of snow mold to growth at temperatures near 0°C has generally allowed them to escape microbial competition in the nival (marked by perpetual snow) environment (19).
Snow cover insulates plants from exposure to lethal and sub-lethal temperatures in winter. Variation in the depth of snow cover creates different climate zones. These attract different snow mold species (because of their susceptibility to temperature), which interact and compete with one another for plant nutrients. Sometimes they are compatible; on other occasions they are incompatible. This highly dynamic system is quite difficult to control.
In northern countries and high elevation regions, application of fungicide is costly and often ineffective for snow mold control. Because chemical reactions are less efficient in cold soils, higher dosages are required to achieve a degree of control comparable to that in other regions. The long, harsh winters also demand a greater degree of persistence than is possessed by most chemical fungicides.
Degradation of chemicals in cold soils is very slow. Because of the fragility of the environment in which snow molds occur, use of chemicals also elicits great concern. For instance, in Alaska, three years after initial application, detectable residues of both metribuzin and triallate (2 and 14%, respectively, of that applied) remained in the soil (6).
Biological agents such as Trichoderma atroviride have been found to possess the persistence and specificity required for control most of the snow mold fungi. Biocontrol promises benefits to agriculture and landscaping, especially in the north, without producing adverse environmental consequences.
Acknowledgments
Research on snow molds and their biological control is part of two Hatch projects supported through the Agricultural and Forestry Experiment Station at the University of Alaska Fairbanks. I thank Mitch Adelman for his technical support; Takako Yokogi, Charity Gitschel and Dr. Peter Gay for their assistance in the preparation of this manuscript. I thank Mr. Joe Holty for the use of his photo of chlorophyll leaching into the snow. I would also like to thank Dr. Jerry A. McBeath and Dr. Gay for critically reviewing the manuscript.
Additional Resources
Snow Mold, Pink or Fusarium and Microdochium patch from Agriculture, Food and Rural Development of Alberta
Cottony Snow Mold or LTB from Agriculture, Food and Rural Development of Alberta
Horticulture & Home Pest News, Iowa State University
Yard & Garden News, University of Minnesota Extension
Snow Molds Damage Lawns and Golf Courses, University of Wisconsin Extension
Managing Turfgrass Diseases, Penn State University
Diagnosing Pink and Gray Snow Molds, University of Guelph
Diseases of Wheat, International Society for Molecular Plant-Microbe Interactions
Snow Mold, Colorado State University Cooperative Extension
References
1. Amano, Y., and Ozeki, S. 1981. Winter wheat breeding for resistance to snow mold and cold hardiness. I. Development of testing methods and application for the classification of resistant varieties. Bull. Hokkaido Pref. Agric. Exp. Stn. 46:12-21.
2. Bazzingher, G. 1976. Der schwarze Schneeschimmel der Koniferen [Herpotrichia juniperi (Duby) Petrak und Herpotrichia coulteri (Peck) Bose]. Eu. J. For. Pathol. 6:109-122.
3. Bruehl, G. W. 1982. Developing wheat resistant to snow mold in Washington state. Plant Dis. 66:1091-1095.
4. Burpee, L. L., Kaye, L. M., Goulty, L. G., and Lawton, M. B. 1987. Suppression of gray snow mold on creeping bentgrass by an isolate of Typhula phacorrhiza. Plant Dis. 71:97-100.
5. Cheng, M., Gay, P. A., and McBeath, J. H. 2001. Determination of chitinolytic activity in under differing environmental conditions. P. 57-62. IN: Proceeings of Biocontrol in a New Millenium: Building for the Future on Past Experience. D. M. Huber, ed.
6. Conn, J., and Cameron, J. 1988. Persistence and carry-over of metribuzin and triallate in subarctic soils. Can. J. Soil Sci. 68:827-837.
7. Conners, I. L. 1967. Annotated index of plant diseases in Canada. Research Branch Canada Depart. Of Agriculture. 381pp.
8. Elenkin, A. A. 1911.On fungal diseases of tulip bulbs. Plant Protect. 5:105-124 (in Russ.)
9. Gaudet, D. A. 1994. Progress towards understanding interactions betweem cold hardiness and snow mold resistance and development of resistant cultivars. Can. J. Plant Pathol. 16:241-146.
10. Gaudet, D. A., and Laroche, A. 1999. Snow mold crop-environment interactions. P. 191- IN: Biotechnological Applications of Cold-Adapted Organisms. Margesin, R. and Schinner, R. eds. Springer-Verlag. 338pp.
11. Gaudet, D. A., Laroche, A., and Yoshida, M. 1999. Low temperqture-wheat-fungal interactions: A carbohydrate connection. Physiological Planarum 106:437-444.
12. Gaudet, D. A., and Scholberg, P. L. 1990. Comparative pathogenicity of Coprinus psychromorbidus monokaryons and dikaryons on winter wheat, alfalfa, grasses, and pome fruit. Can. J. Plant Pathol. 12:31-37.
13. Hsiang, T., Matsumoto, N., Millett, S. M. 1999. Biology and Management of Typhula snow molds of turfgrass. Plant Dis. 83:783-798.
14. Istokovics, A., Morita, N., Isumi, K., Hoshino, T., Yumoto, I., Sawada, M. T., Ishizaki, K., and Okuyama, H. 1998. Nutral lipids, phospholipids and a betaine lipid of the snow mold fungus Microdochium nivale. Can. J. Microbiol. 44:1051-1059.
15. Kiyomoto, R. K., and Bruehl, G. W. 1977. Carbohydrate accumulation and depletion by winter cereals differing in resistance to Typhula idahoensis. Phytopathology 67:206-211.
16. Kurkela, T. T. 1996. Ascospore production period of Phacidium infestans, a snow blight fungus on Pinus sylvestris. Scand. J. For. Res. 11:60-67.
17. Labeau, J. B. 1975. Antagonisms between isolates of a snow mold pathogen. 65:877-880.
18. Matsumoto, N. 1994. Ecological adaptations of low temperature plant pathogenic fungi to diverse winter climates. Can. J. Plant Pathol. 16:237-240.
19. Matsumoto, N., and Tajimi, A. 1987. Bacterial flora associated with the snow mold fungi, Typhula incarnata and T. ishikariensis. Ann. Phytopath. Soc. Japan 53:250-253.
20. Matsumoto, N., and Tajimi, A. 1993. Effect of cropping history on the population structure of Typhula incarnata and Typhula ishikariensis. Can. J. Bot. 71:143401440.
21. McBeath, J. H. 2001. Effects of Trichoderma atroviride on snow mold diseases of turfgrass in interior Alaska. P. 98-101. IN: Proceeings of Biocontrol in a New Millenium: Building for the Future on Past Experience. Huber, D.M. ed.
22. McBeath, J. H. 1985. Pink snow mold on winter cereals and lawn grasses in Alaska. Plant Dis. 69:722-723.
23. McBeath, J. H. 1988. Sclerotial myceliogenic germination and pathogenicity of Sclerotinia borealis. IN: Proceedings of the 5th International Congress of Plant Pathology. p.201.
24. McBeath, J. H., and Adelman, M. 1987. Cyanogenesis of a sclerotial low temperature basidiomycete. Phytopathology 77:1701.
25. McBeath, J. H., and Wenko, L. 1986. A simple, versatile method to determine extracellular enzymes in snow mold. Phytopathology 76:1143.
26. McBeath, J. H., Mehdizadegen, F., and Lackwood, H. 1990. Anther culture selection to enhance snow mold disease resistance in winter wheat. Pages 258-263 in: Progress in Plant Cellular and Molecular Biology. Current Plant Science and Biotechnology in Agriculture. H. J. J. Nijkamp, L. H. W. Van Der Plas, and J. Van Aartrijk, eds. Kluwer Academic Publishers.
27. Morita, R. Y. 1975. Psychrophilic bacteria. Bacteril. Rev. 39:144-167.
28. Saito, I. 1998. Non-gramineous hosts of Myriosclerotinia borealis. Mycoscience 39:146-163.
29. Smiley, R. W., Dernoeden, P. H., and Clarke, B. B. 1992. Compendium of Turfgrass Diseases. 2nd ed. APS Press.
30. Simms. H. R. 1967. On the ecology of Herpotrichia nigra. Mycologia 59:902-909.
31. Smith, J. D. 1987. Winter-hardiness and overwintering diseases of amenity turfgrasses with special references to the Canadian prairies. Can. Tech. Bull. 1987-12E.
32. Smith, J. D., and Davidson, J. G. N. 1979. Acremonium boreale n.sp., a sclerotial, low-temperature-tolerant, snow mold antagonist. Can. J. Bot. 57:2122-2139.
33. Smith, J. D., Jackson, N., and Woolhouse, A. R. 1989. Fungal Diseases of Amenity Turf Grasses. F. & F. N. Spon, London
34. Sunderman, D. W. 1964. Modifications of the Cormack and Lebeau technique for inoculated winter wheat with snow-mold causing Typhula species. Plant Dis. Rep. 48:394-395.
35. Takasawa, T., Sagisaka, K., Yagi, K., Uchiyama, K., Aoki, A., Takaoka, K., and Yamamato, K. 1997. Polygalacturonase isolated from the culture of the psychrophilic fungus Sclerotinia borealis. Can. J. Microbiol. 43:417-424.
36. Takenaka, S., and Arai, M. 1993. Dynamics of three snow mold pathogens Pythium paddicum, Pythium iwayamai, and Typhula incarnata on barley plant tissues. Can. J. Bot. 71:757-763.
37. Tomiyama, K. 1955. Studies on the Snow Blight Disease of Winter Cereals. Hokkaido Nat. Agric. Exp. Stn. Rep. 47. 234 pp.
38. Wong, P. T. W., and McBeath, J. H. 1999. Plant protection by cold-adapted fungi. p. 177- IN: Biotechnological Applications of Cold-Adapted Organisms. Margesin, R. and Schinner, R. eds. Springer-Verlag. 338pp.
39. Wooding, F. J., and McBeath, J. H. 1984. Growing winter grains in Alaska. Agroborealis 16:21-26.
40. Wu, C., Hsinang, T., Yang, I., and Liu, L. X. 1996. Evaluation of Typhula phacorrhiza for the biocontrol of grey snow mold in turfgrass. Pages 227-233 in: Advances in Biological Control of Plant Diseases. W. Tang, R. J. Cook, and A. D. Rovira, eds. Beijing. China Agricultural University Press.
41. Yoshida, M., Abe, J., Moriyama, M., and Kuwabara, T. 1997. Carbohydrate levels among winter wheat cultivars varying in freezing tolerance and snow mold resistance during autumn and winter. Physiologia Plantarum 103:8-16.

